

Role of PRY-1/Axin in heterochronic miRNA-mediated seam cell development
- PMID: 31307392
- PMCID: PMC6631683
- DOI: 10.1186/s12861-019-0197-5
Role of PRY-1/Axin in heterochronic miRNA-mediated seam cell development
Abstract
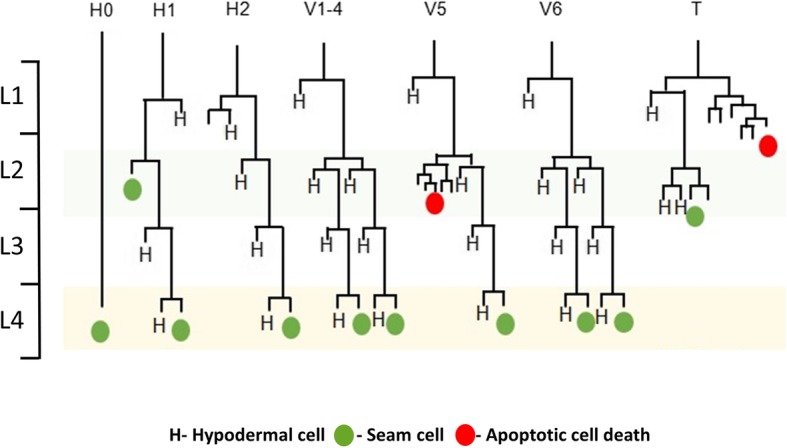
Background: Caenorhabditis elegans seam cells serve as a good model to understand how genes and signaling pathways interact to control asymmetric cell fates. The stage-specific pattern of seam cell division is coordinated by a genetic network that includes WNT asymmetry pathway components WRM-1, LIT-1, and POP-1, as well as heterochronic microRNAs (miRNAs) and their downstream targets. Mutations in pry-1, a negative regulator of WNT signaling that belongs to the Axin family, were shown to cause seam cell defects; however, the mechanism of PRY-1 action and its interactions with miRNAs remain unclear.
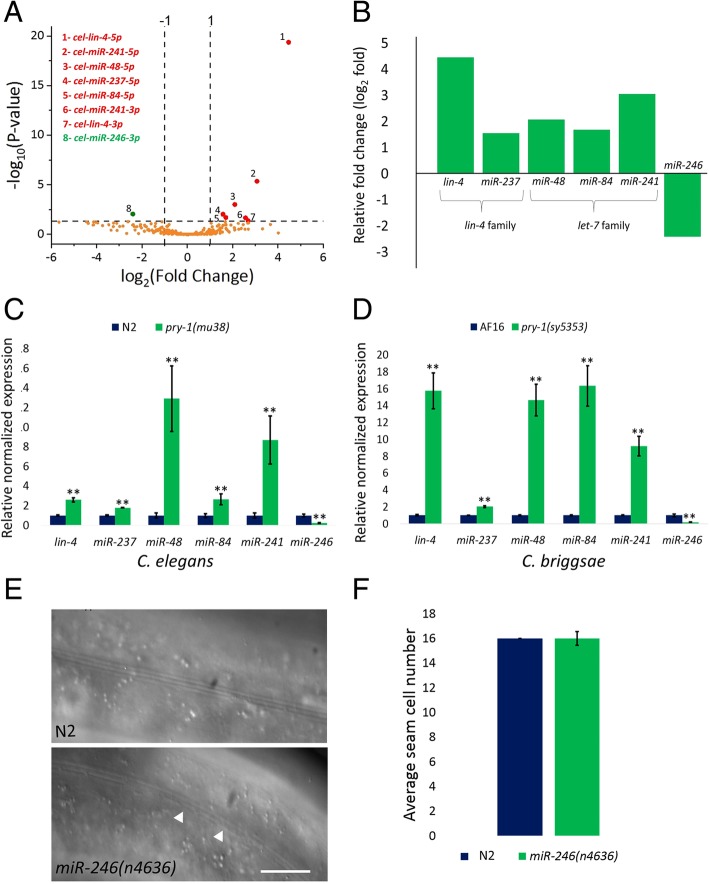
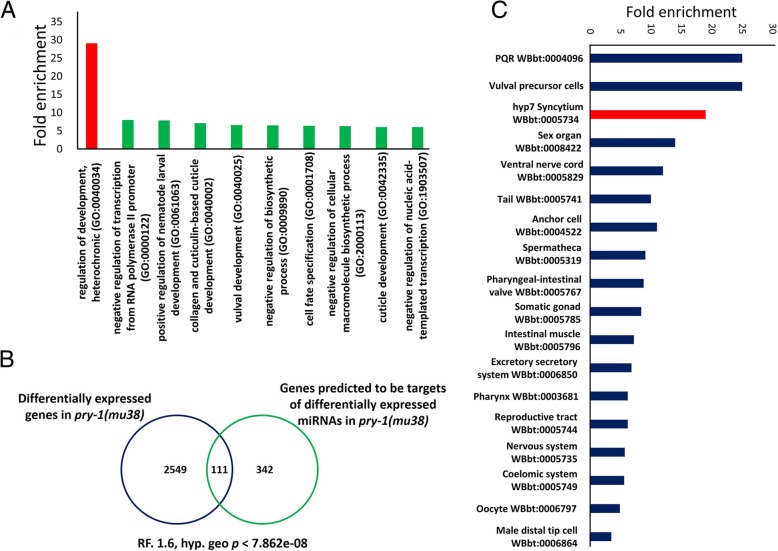
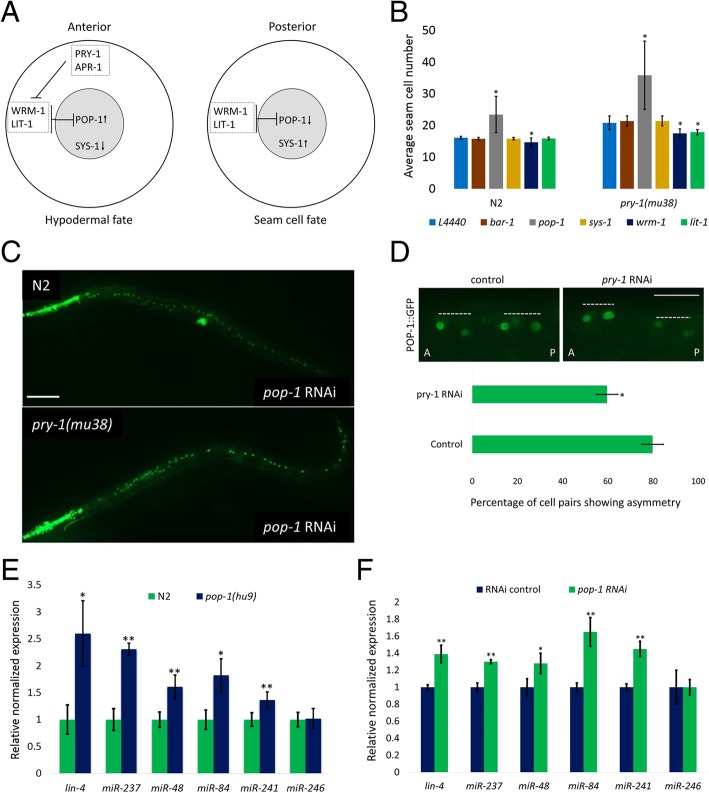
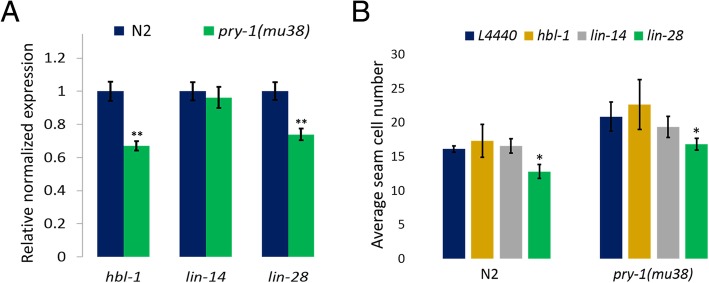
Results: We found that pry-1 mutants in C. elegans exhibit seam cell, cuticle, and alae defects. To examine this further, a miRNA transcriptome analysis was carried out, which showed that let-7 (miR-48, miR-84, miR-241) and lin-4 (lin-4, miR-237) family members were upregulated in the absence of pry-1 function. Similar phenotypes and patterns of miRNA overexpression were also observed in C. briggsae pry-1 mutants, a species that is closely related to C. elegans. RNA interference-mediated silencing of wrm-1 and lit-1 in the C. elegans pry-1 mutants rescued the seam cell defect, whereas pop-1 silencing enhanced the phenotype, suggesting that all three proteins are likely important for PRY-1 function in seam cells. We also found that these miRNAs were overexpressed in pop-1 hypomorphic animals, suggesting that PRY-1 may be required for POP-1-mediated miRNA suppression. Analysis of the let-7 and lin-4-family heterochronic targets, lin-28 and hbl-1, showed that both genes were significantly downregulated in pry-1 mutants, and furthermore, lin-28 silencing reduced the number of seam cells in mutant animals.
Conclusions: Our results show that PRY-1 plays a conserved role to maintain normal expression of heterochronic miRNAs in nematodes. Furthermore, we demonstrated that PRY-1 acts upstream of the WNT asymmetry pathway components WRM-1, LIT-1, and POP-1, and miRNA target genes in seam cell development.
Keywords: Axin; C. briggsae; C. elegans; Heterochronic development; Pry-1; Seam cell; WNT asymmetry pathway; miRNA.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


Similar articles
-
Wnt signaling controls temporal identities of seam cells in Caenorhabditis elegans.Dev Biol. 2010 Sep 15;345(2):144-55. doi: 10.1016/j.ydbio.2010.07.002. Epub 2010 Jul 17. Dev Biol. 2010. PMID: 20624379
-
acn-1, a C. elegans homologue of ACE, genetically interacts with the let-7 microRNA and other heterochronic genes.Cell Cycle. 2017 Oct 2;16(19):1800-1809. doi: 10.1080/15384101.2017.1344798. Epub 2017 Sep 21. Cell Cycle. 2017. PMID: 28933985 Free PMC article.
-
Wnt signaling controls the stem cell-like asymmetric division of the epithelial seam cells during C. elegans larval development.Dev Biol. 2010 Dec 1;348(1):58-66. doi: 10.1016/j.ydbio.2010.09.005. Epub 2010 Sep 16. Dev Biol. 2010. PMID: 20849842 Free PMC article.
-
Heterochronic control of AFF-1-mediated cell-to-cell fusion in C. elegans.Adv Exp Med Biol. 2011;713:5-11. doi: 10.1007/978-94-007-0763-4_2. Adv Exp Med Biol. 2011. PMID: 21432011 Review.
-
An elegant miRror: microRNAs in stem cells, developmental timing and cancer.Chromosoma. 2009 Aug;118(4):405-18. doi: 10.1007/s00412-009-0210-z. Epub 2009 Apr 3. Chromosoma. 2009. PMID: 19340450 Free PMC article. Review.
Cited by
-
Mechanisms of lineage specification in Caenorhabditis elegans.Genetics. 2023 Dec 6;225(4):iyad174. doi: 10.1093/genetics/iyad174. Genetics. 2023. PMID: 37847877 Free PMC article.
-
EFL-3/E2F7 modulates Wnt signalling by repressing the Nemo-like kinase LIT-1 during asymmetric epidermal cell division in Caenorhabditis elegans.Development. 2025 Mar 1;152(5):DEV204546. doi: 10.1242/dev.204546. Epub 2025 Mar 3. Development. 2025. PMID: 40026193 Free PMC article.
-
Axin Family of Scaffolding Proteins in Development: Lessons from C. elegans.J Dev Biol. 2019 Oct 15;7(4):20. doi: 10.3390/jdb7040020. J Dev Biol. 2019. PMID: 31618970 Free PMC article. Review.
-
AXIN-AMPK signaling: Implications for healthy aging.F1000Res. 2021 Dec 8;10:1259. doi: 10.12688/f1000research.74220.1. eCollection 2021. F1000Res. 2021. PMID: 35087668 Free PMC article. Review.
-
Genetic analysis of Caenorhabditis elegans pry-1/Axin suppressors identifies genes involved in reproductive structure development, stress responses, and aging.G3 (Bethesda). 2022 Feb 4;12(2):jkab430. doi: 10.1093/g3journal/jkab430. G3 (Bethesda). 2022. PMID: 35100345 Free PMC article.
References
-
- Hall DH, Altun ZF. C. elegans Atlas. New York: Cold Spring Harbor Laboratory Press; 2008.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
