

Canine platelets express functional Toll-like receptor-4: lipopolysaccharide-triggered platelet activation is dependent on adenosine diphosphate and thromboxane A2 in dogs
- PMID: 31307465
- PMCID: PMC6632210
- DOI: 10.1186/s12917-019-1997-3
Canine platelets express functional Toll-like receptor-4: lipopolysaccharide-triggered platelet activation is dependent on adenosine diphosphate and thromboxane A2 in dogs
Abstract
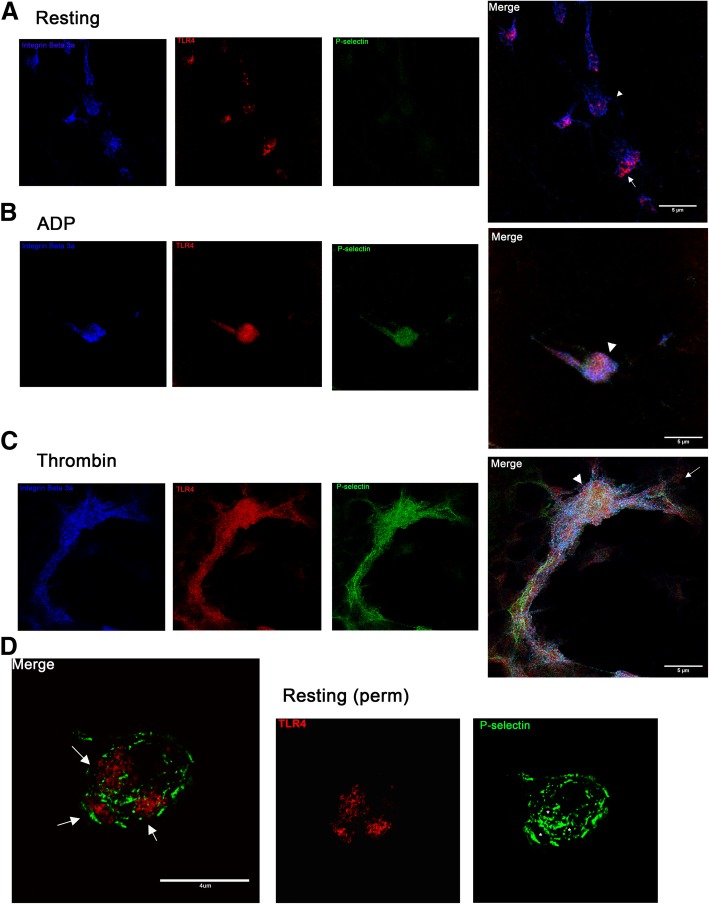
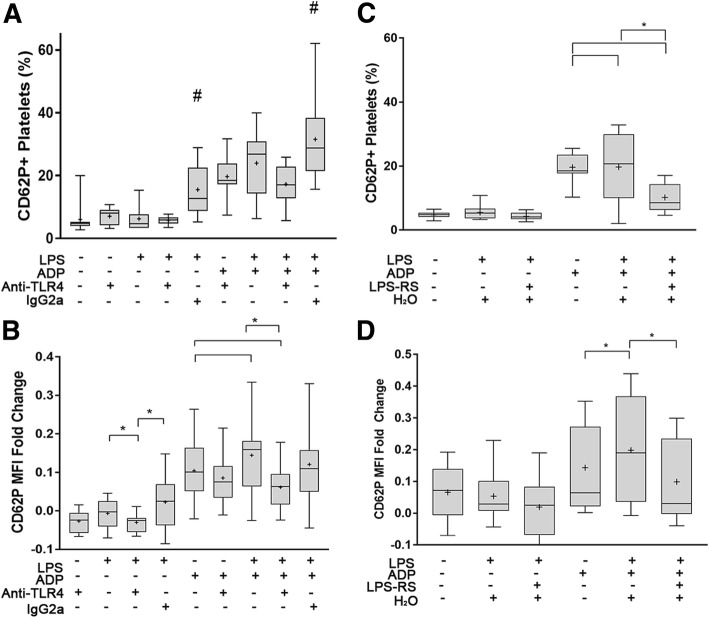
Background: Functional Toll-like receptor 4 (TLR4) has been characterized in human and murine platelets indicating that platelets play a role in inflammation and hemostasis during sepsis. It is unclear whether canine platelets could express functional TLR4 by responding to its ligand, lipopolysaccharide (LPS). We sought to determine if dogs express functional TLR4 and if LPS-induced platelet activation requires co-stimulation with ADP or thromboxane A2 (TxA2). Canine platelets were unstimulated (resting) or activated with thrombin or ADP prior to flow cytometric or microscopic analyses for TLR4 expression. We treated resting or ADP-primed platelets with LPS in the absence or presence of acetylsalicylic acid (ASA) and inhibited TLR4 with function blocking antibody or LPS from Rhodobacter sphaeroides (LPS-RS).
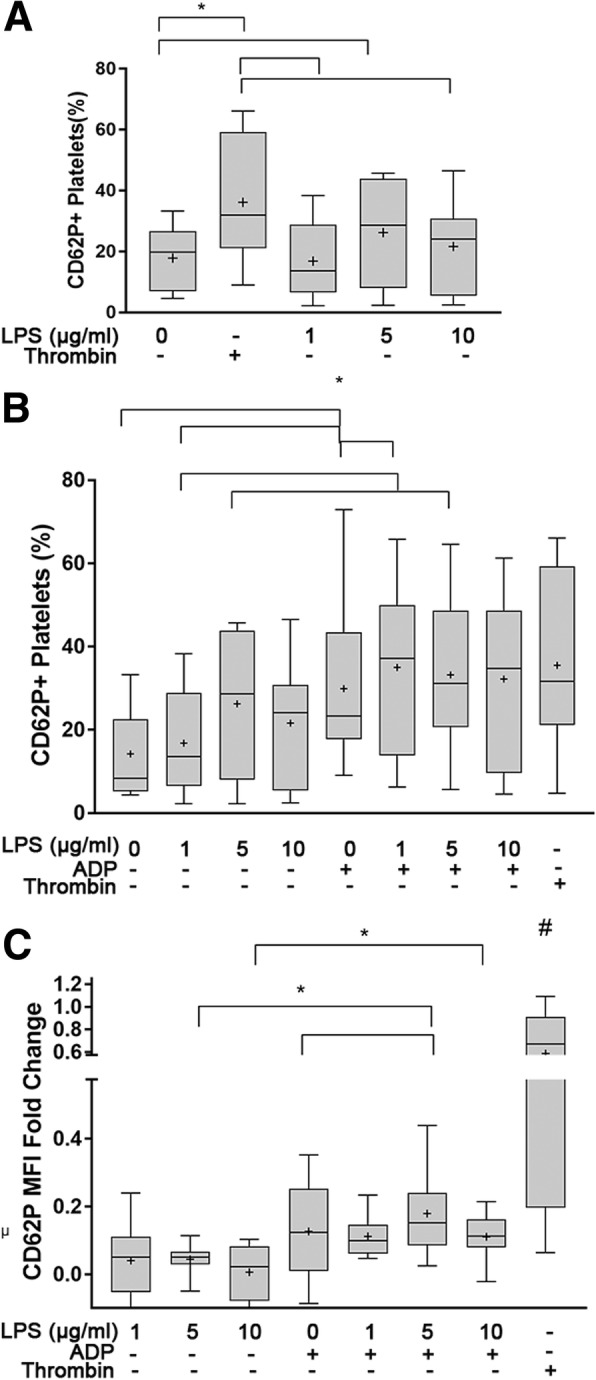
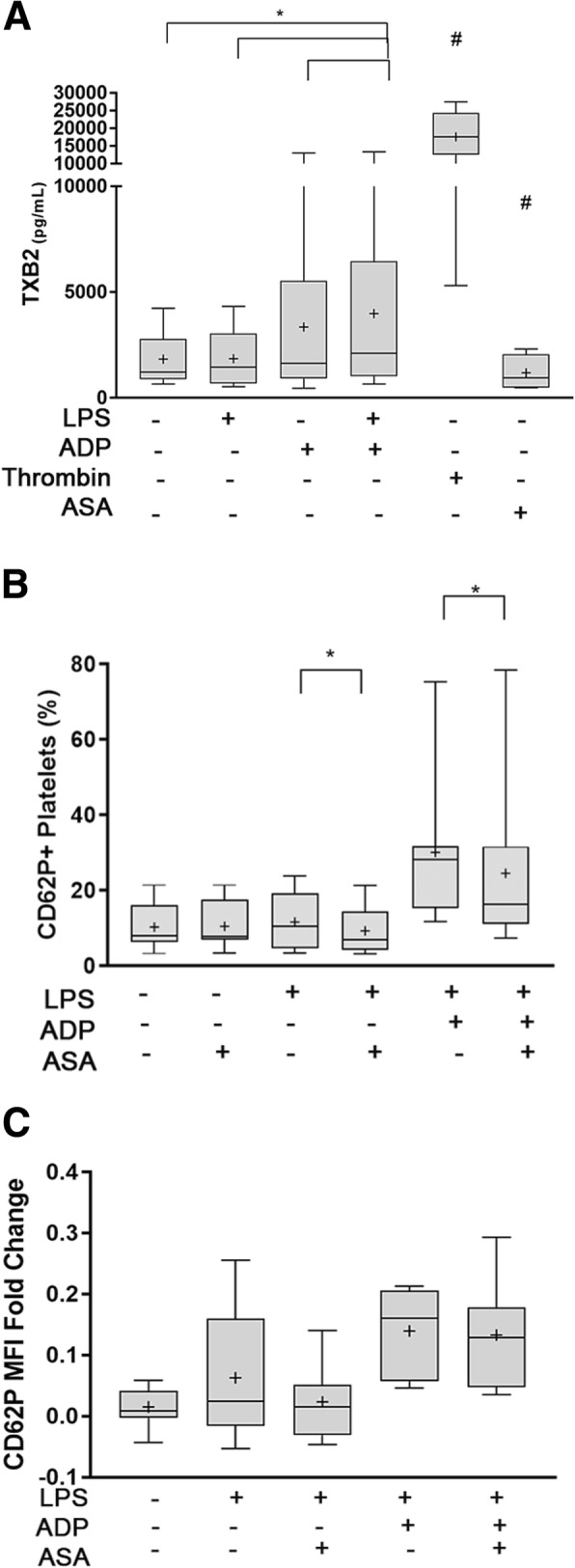
Results: We discovered that dog platelets have variable TLR4 expression, which was upregulated following thrombin or ADP activation. LPS augmented P-selectin expression and thromboxane B2 secretion in ADP-primed platelets via TLR4. Inhibition of cyclooxygenase by ASA attenuated LPS-mediated P-selectin expression demonstrating that TLR4 signaling in platelets is partially dependent on TxA2 pathway.
Conclusion: Expression of functional TLR4 on canine platelets may contribute to hypercoagulability in clinical septic dogs. Cyclooxygenase and TxA2 pathways in TLR4-mediated platelet activation may present novel therapeutic targets in dogs with sepsis.
Keywords: Aspirin; Inflammation; Pattern recognition receptor; Platelet-priming; Sepsis; Thrombosis.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
-
- Duerschmied D, Bode C, Ahrens I. Immune functions of platelets. Thromb Haemost. 2014;112(4):678–691. - PubMed
