

Sustained Oscillations of Epithelial Cell Sheets
- PMID: 31307676
- PMCID: PMC6697349
- DOI: 10.1016/j.bpj.2019.06.013
Sustained Oscillations of Epithelial Cell Sheets
Abstract
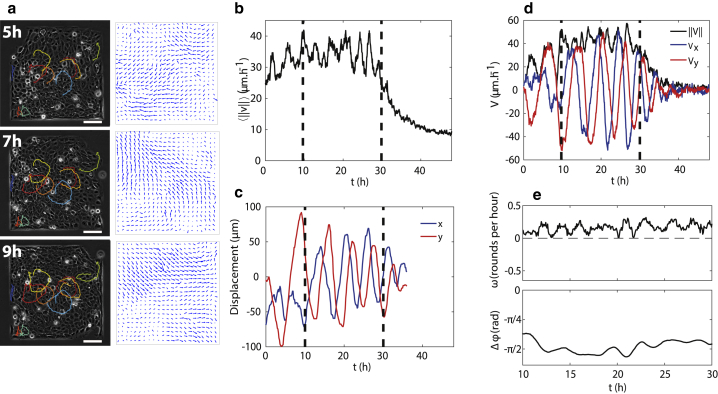
Morphological changes during development, tissue repair, and disease largely rely on coordinated cell movements and are controlled by the tissue environment. Epithelial cell sheets are often subjected to large-scale deformation during tissue formation. The active mechanical environment in which epithelial cells operate have the ability to promote collective oscillations, but how these cellular movements are generated and relate to collective migration remains unclear. Here, combining in vitro experiments and computational modeling, we describe a form of collective oscillations in confined epithelial tissues in which the oscillatory motion is the dominant contribution to the cellular movements. We show that epithelial cells exhibit large-scale coherent oscillations when constrained within micropatterns of varying shapes and sizes and that their period and amplitude are set by the smallest confinement dimension. Using molecular perturbations, we then demonstrate that force transmission at cell-cell junctions and its coupling to cell polarity are pivotal for the generation of these collective movements. We find that the resulting tissue deformations are sufficient to trigger osillatory mechanotransduction of YAP within cells, potentially affecting a wide range of cellular processes.
Copyright © 2019 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures





Similar articles
-
Force localization modes in dynamic epithelial colonies.Mol Biol Cell. 2018 Nov 15;29(23):2835-2847. doi: 10.1091/mbc.E18-05-0336. Epub 2018 Sep 12. Mol Biol Cell. 2018. PMID: 30207837 Free PMC article.
-
Guidance of collective cell migration by substrate geometry.Integr Biol (Camb). 2013 Aug;5(8):1026-35. doi: 10.1039/c3ib40054a. Integr Biol (Camb). 2013. PMID: 23784144
-
Regulation of epithelial cell organization by tuning cell-substrate adhesion.Integr Biol (Camb). 2015 Oct;7(10):1228-41. doi: 10.1039/c5ib00196j. Epub 2015 Sep 24. Integr Biol (Camb). 2015. PMID: 26402903 Free PMC article.
-
Phenomenological approaches to collective behavior in epithelial cell migration.Biochim Biophys Acta. 2015 Nov;1853(11 Pt B):3143-52. doi: 10.1016/j.bbamcr.2015.05.021. Epub 2015 May 29. Biochim Biophys Acta. 2015. PMID: 26028592 Review.
-
Cadherin mechanotransduction in leader-follower cell specification during collective migration.Exp Cell Res. 2019 Mar 1;376(1):86-91. doi: 10.1016/j.yexcr.2019.01.006. Epub 2019 Jan 8. Exp Cell Res. 2019. PMID: 30633881 Review.
Cited by
-
Force propagation between epithelial cells depends on active coupling and mechano-structural polarization.Elife. 2023 Aug 7;12:e83588. doi: 10.7554/eLife.83588. Elife. 2023. PMID: 37548995 Free PMC article.
-
Engineering tools for quantifying and manipulating forces in epithelia.Biophys Rev (Melville). 2023 May 11;4(2):021303. doi: 10.1063/5.0142537. eCollection 2023 Jun. Biophys Rev (Melville). 2023. PMID: 38510344 Free PMC article. Review.
-
Cell migration guided by long-lived spatial memory.Nat Commun. 2021 Jul 5;12(1):4118. doi: 10.1038/s41467-021-24249-8. Nat Commun. 2021. PMID: 34226542 Free PMC article.
-
In vivo engraftment into the cornea endothelium using extracellular matrix shrink-wrapped cells.Commun Mater. 2022;3(1):25. doi: 10.1038/s43246-022-00247-1. Epub 2022 Apr 26. Commun Mater. 2022. PMID: 39175945 Free PMC article.
-
Picking winners in cell-cell collisions: Wetting, speed, and contact.Phys Rev E. 2022 Nov;106(5-1):054413. doi: 10.1103/PhysRevE.106.054413. Phys Rev E. 2022. PMID: 36559372 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
