

APE1 Upregulates MMP-14 via Redox-Sensitive ARF6-Mediated Recycling to Promote Cell Invasion of Esophageal Adenocarcinoma
- PMID: 31308045
- PMCID: PMC6726537
- DOI: 10.1158/0008-5472.CAN-19-0237
APE1 Upregulates MMP-14 via Redox-Sensitive ARF6-Mediated Recycling to Promote Cell Invasion of Esophageal Adenocarcinoma
Abstract
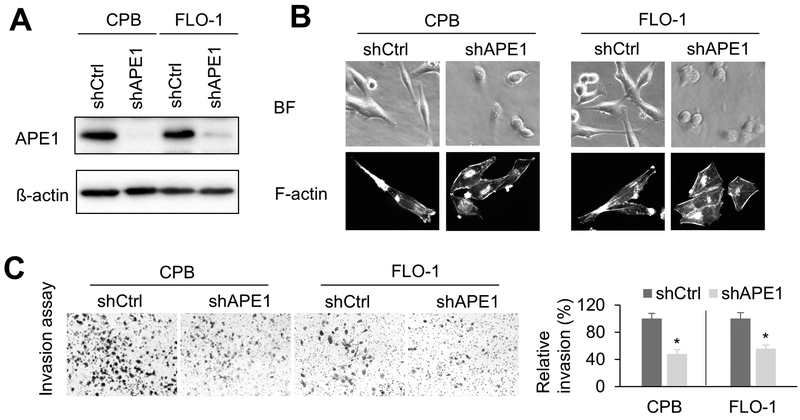
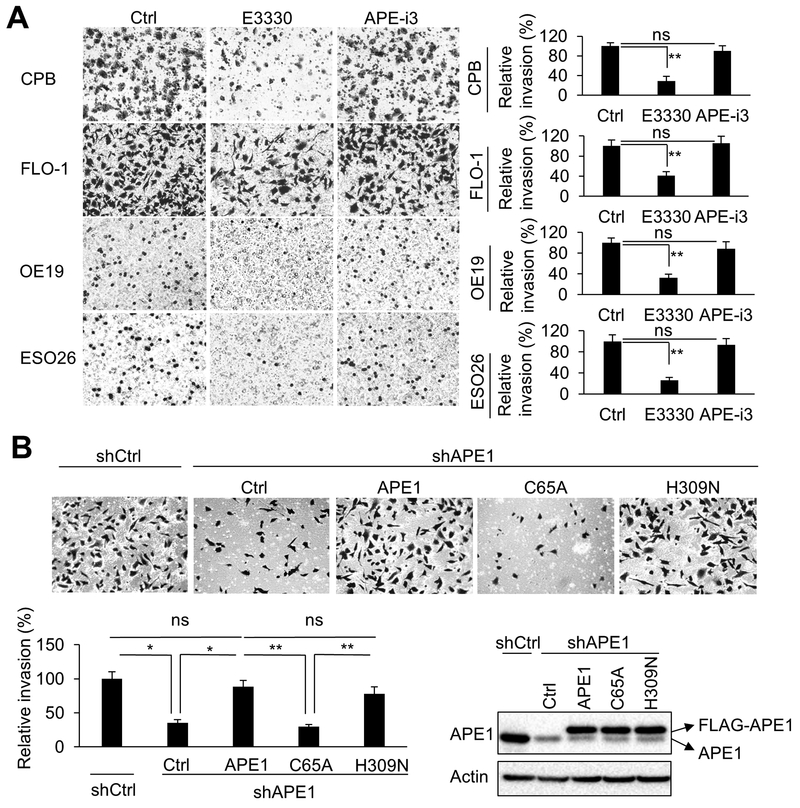
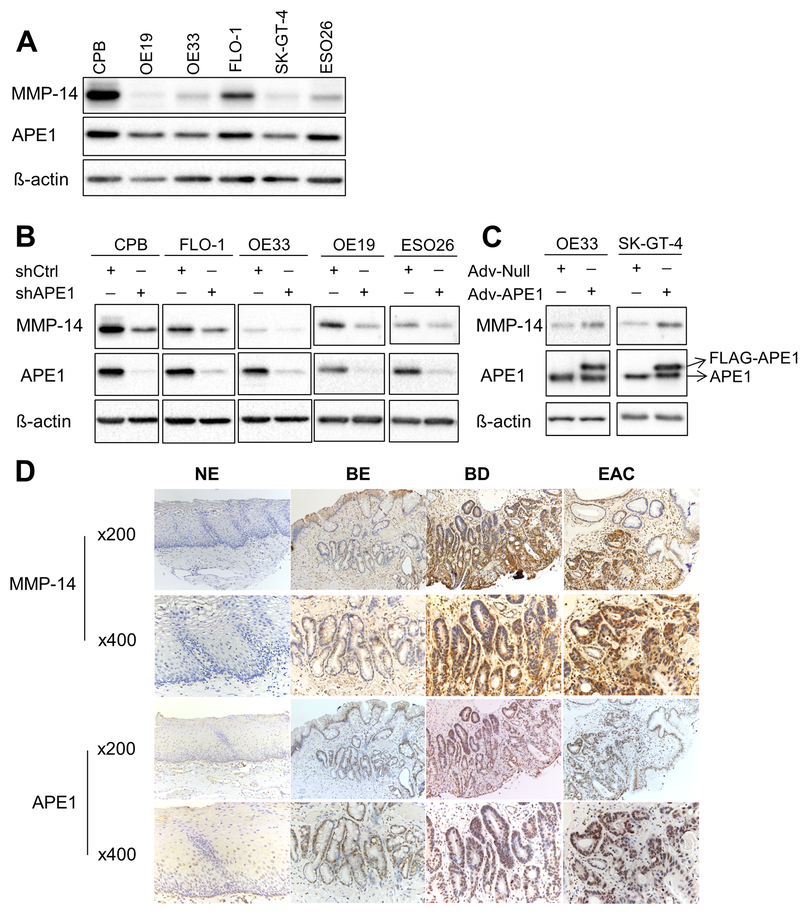
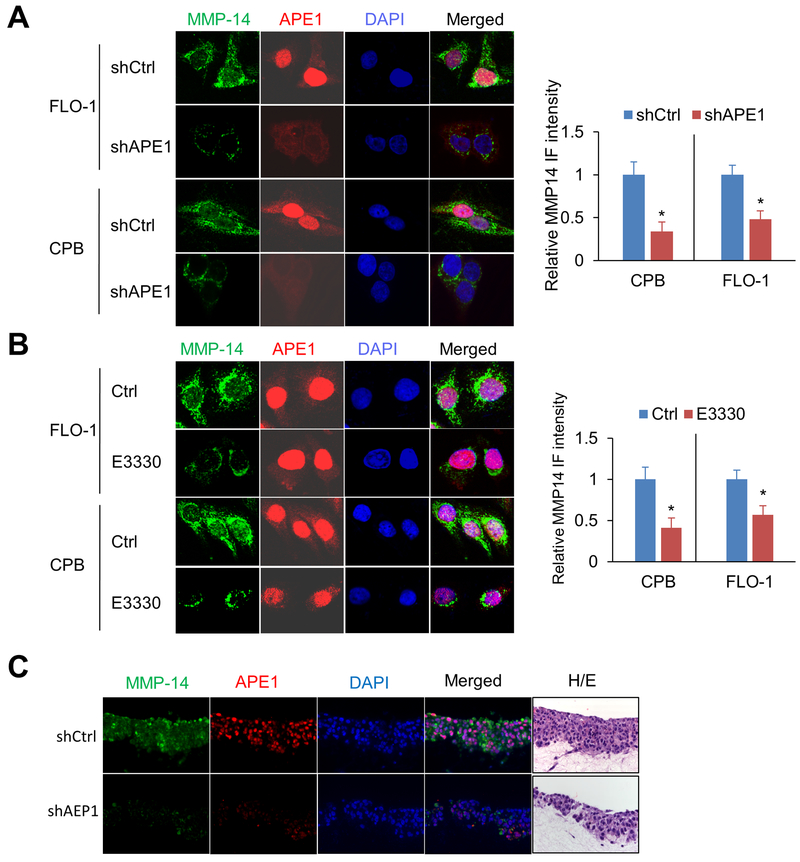
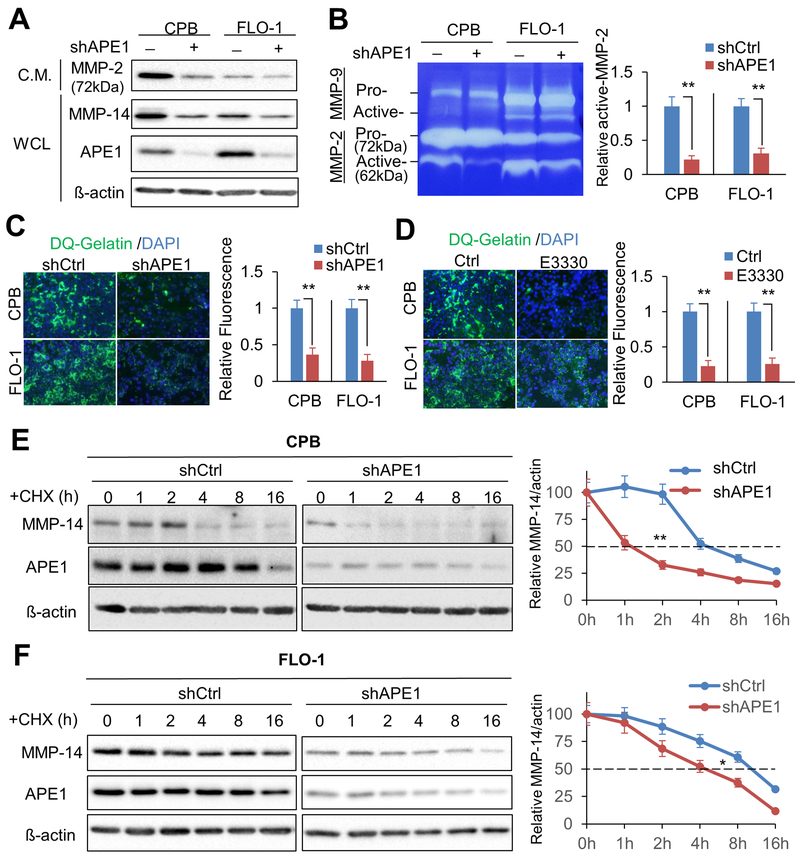
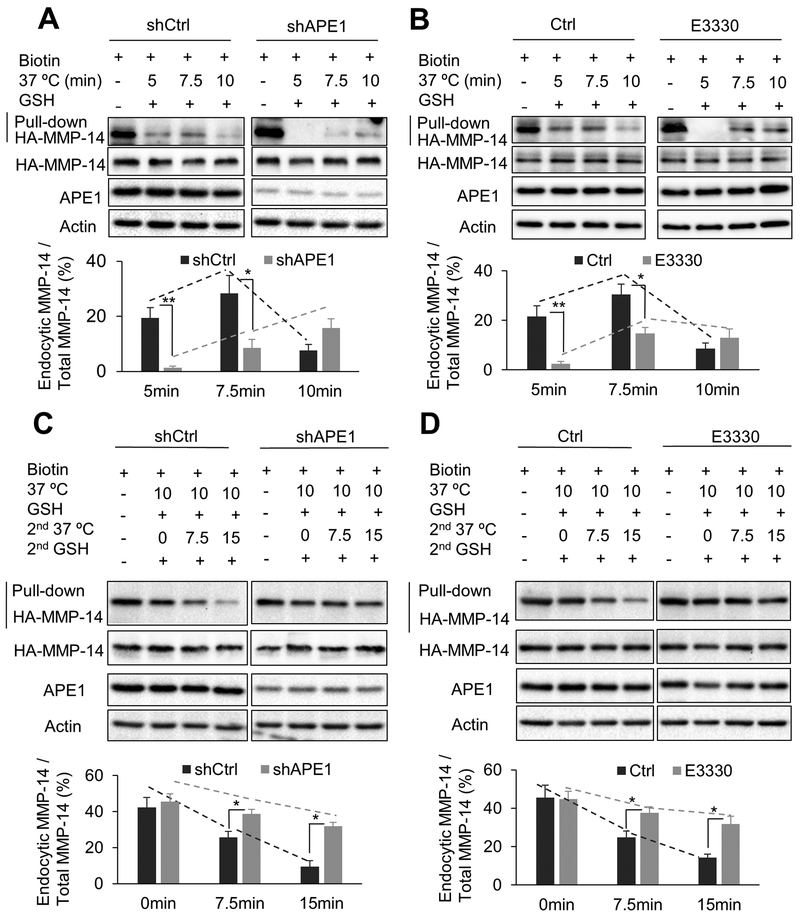
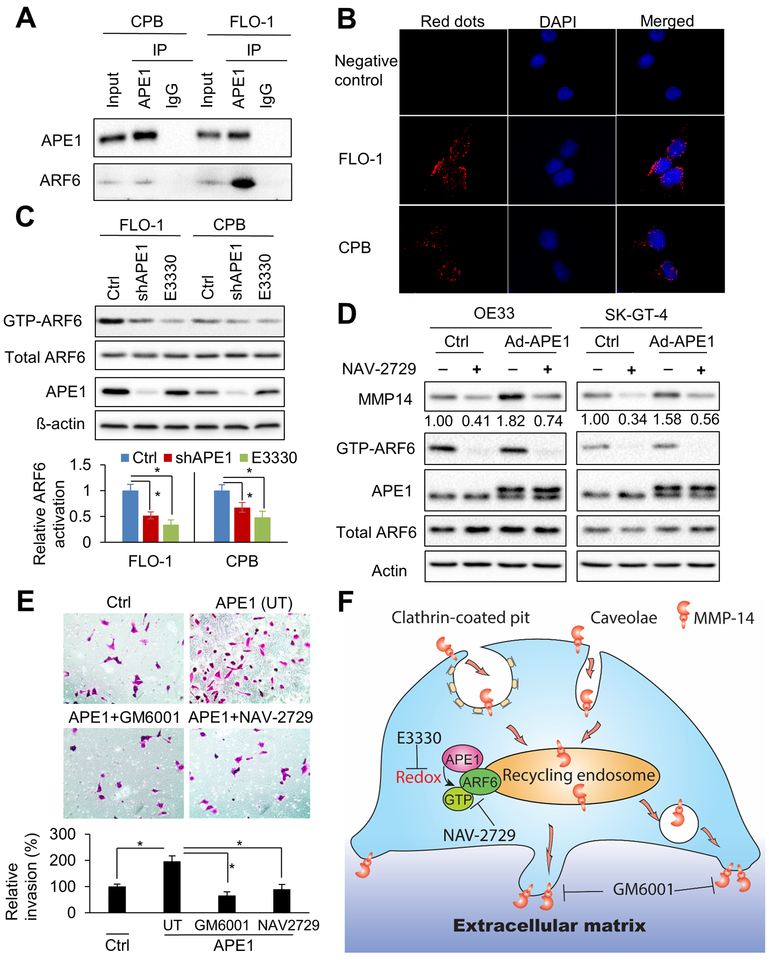
Esophageal adenocarcinoma (EAC) is an aggressive malignancy with poor clinical outcome. The incidence of EAC has been rising rapidly in the past three decades. Here, we showed that apurinic/apyrimidinic endonuclease (APE1) is overexpressed in EAC cell lines, and patients' samples of dysplasia and EAC. Downregulation of APE1 or inhibition of its redox function significantly repressed invasion. Overexpression of a redox-defective mutant, C65A, abrogated the proinvasive phenotype of APE1. APE1 regulated invasion via upregulation of matrix metalloproteinase 14 (MMP-14), which subsequently activated MMP-2, leading to degradation of the extracellular matrix in a redox-dependent manner. Downregulation of APE1 or inhibition of its redox function decreased the rate of endocytosis and recycling of MMP-14 protein. APE1 interacted with ARF6, a key regulator of MMP-14 recycling, which maintained ARF6 activity in an APE1-redox-dependent manner, promoting its ability to regulate MMP-14 recycling to the cell surface. In summary, these findings identify a novel redox-sensitive APE1-ARF6-MMP-14 signaling axis that mediates cellular invasion in esophageal carcinogenesis. SIGNIFICANCE: This study demonstrates the association between oxidative stress and the development and metastatic behavior of esophageal adenocarcinoma.
©2019 American Association for Cancer Research.
Conflict of interest statement
No potential conflicts of interest were disclosed.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
