

Cytosolic Ca2+ Buffers Are Inherently Ca2+ Signal Modulators
- PMID: 31308146
- PMCID: PMC6942125
- DOI: 10.1101/cshperspect.a035543
Cytosolic Ca2+ Buffers Are Inherently Ca2+ Signal Modulators
Abstract
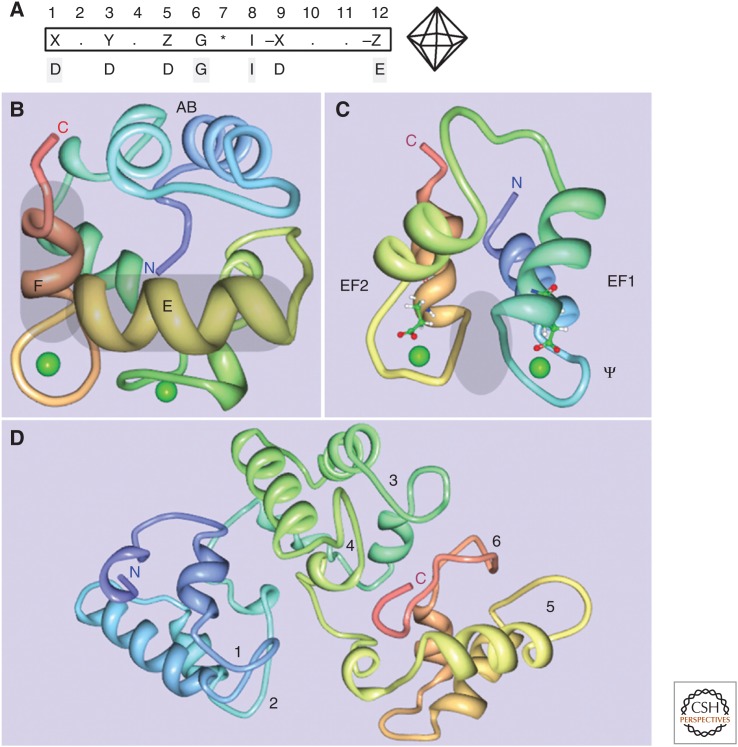
For precisely regulating intracellular Ca2+ signals in a time- and space-dependent manner, cells make use of various components of the "Ca2+ signaling toolkit," including Ca2+ entry and Ca2+ extrusion systems. A class of cytosolic Ca2+-binding proteins termed Ca2+ buffers serves as modulators of such, mostly short-lived Ca2+ signals. Prototypical Ca2+ buffers include parvalbumins (α and β isoforms), calbindin-D9k, calbindin-D28k, and calretinin. Although initially considered to function as pure Ca2+ buffers, that is, as intracellular Ca2+ signal modulators controlling the shape (amplitude, decay, spread) of Ca2+ signals, evidence has accumulated that calbindin-D28k and calretinin have additional Ca2+ sensor functions. These other functions are brought about by direct interactions with target proteins, thereby modulating their targets' function/activity. Dysregulation of Ca2+ buffer expression is associated with several neurologic/neurodevelopmental disorders including autism spectrum disorder (ASD) and schizophrenia. In some cases, the presence of these proteins is presumed to confer a neuroprotective effect, as evidenced in animal models of Parkinson's or Alzheimer's disease.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures

Similar articles
-
Cytosolic Ca2+ buffers.Cold Spring Harb Perspect Biol. 2010 Nov;2(11):a004051. doi: 10.1101/cshperspect.a004051. Epub 2010 Oct 13. Cold Spring Harb Perspect Biol. 2010. PMID: 20943758 Free PMC article. Review.
-
The continuing disappearance of "pure" Ca2+ buffers.Cell Mol Life Sci. 2009 Jan;66(2):275-300. doi: 10.1007/s00018-008-8564-6. Cell Mol Life Sci. 2009. PMID: 19099190 Free PMC article. Review.
-
The use of transgenic mouse models to reveal the functions of Ca2+ buffer proteins in excitable cells.Biochim Biophys Acta. 2012 Aug;1820(8):1294-303. doi: 10.1016/j.bbagen.2011.11.008. Epub 2011 Nov 27. Biochim Biophys Acta. 2012. PMID: 22138448 Review.
-
Calretinin Functions in Malignant Mesothelioma Cells Cannot Be Replaced by the Closely Related Ca2+-Binding Proteins Calbindin-D28k and Parvalbumin.Int J Mol Sci. 2018 Dec 12;19(12):4015. doi: 10.3390/ijms19124015. Int J Mol Sci. 2018. PMID: 30545133 Free PMC article.
-
Calcium buffering properties of calbindin D28k and parvalbumin in rat sensory neurones.J Physiol. 1993 Dec;472:341-57. doi: 10.1113/jphysiol.1993.sp019950. J Physiol. 1993. PMID: 8145149 Free PMC article.
Cited by
-
Stationary Ca2+ nanodomains in the presence of buffers with two binding sites.Biophys J. 2021 May 18;120(10):1942-1956. doi: 10.1016/j.bpj.2021.03.015. Epub 2021 Mar 23. Biophys J. 2021. PMID: 33771472 Free PMC article.
-
Magnetic domains oscillation in the brain with neurodegenerative disease.Sci Rep. 2021 Jan 12;11(1):714. doi: 10.1038/s41598-020-80212-5. Sci Rep. 2021. PMID: 33436793 Free PMC article.
-
Physiology of intracellular calcium buffering.Physiol Rev. 2023 Oct 1;103(4):2767-2845. doi: 10.1152/physrev.00042.2022. Epub 2023 Jun 16. Physiol Rev. 2023. PMID: 37326298 Free PMC article. Review.
-
Calcium Signaling Dynamics in Vascular Cells and Their Dysregulation in Vascular Disease.Biomolecules. 2025 Jun 18;15(6):892. doi: 10.3390/biom15060892. Biomolecules. 2025. PMID: 40563532 Free PMC article. Review.
-
Rabies virus infection is associated with variations in calbindin D-28K and calretinin mRNA expression levels in mouse brain tissue.Arch Virol. 2023 Apr 18;168(5):143. doi: 10.1007/s00705-023-05753-2. Arch Virol. 2023. PMID: 37069450 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous