

Perturbation of ubiquitin homeostasis promotes macrophage oxidative defenses
- PMID: 31308397
- PMCID: PMC6629656
- DOI: 10.1038/s41598-019-46526-9
Perturbation of ubiquitin homeostasis promotes macrophage oxidative defenses
Abstract
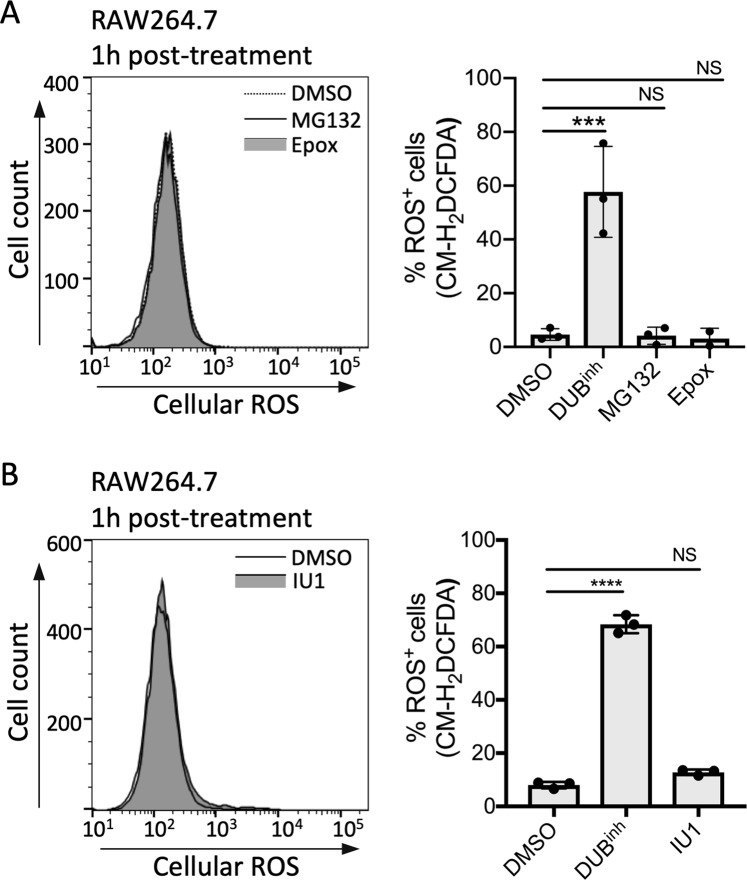
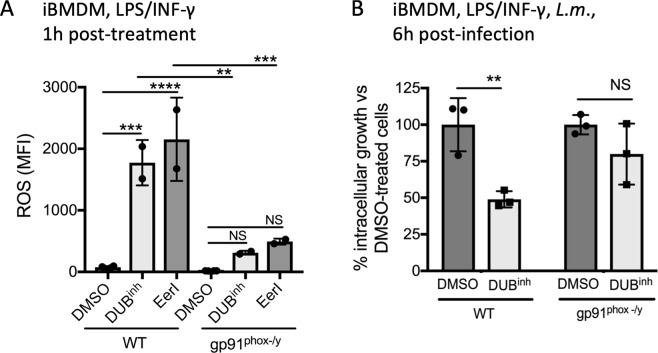
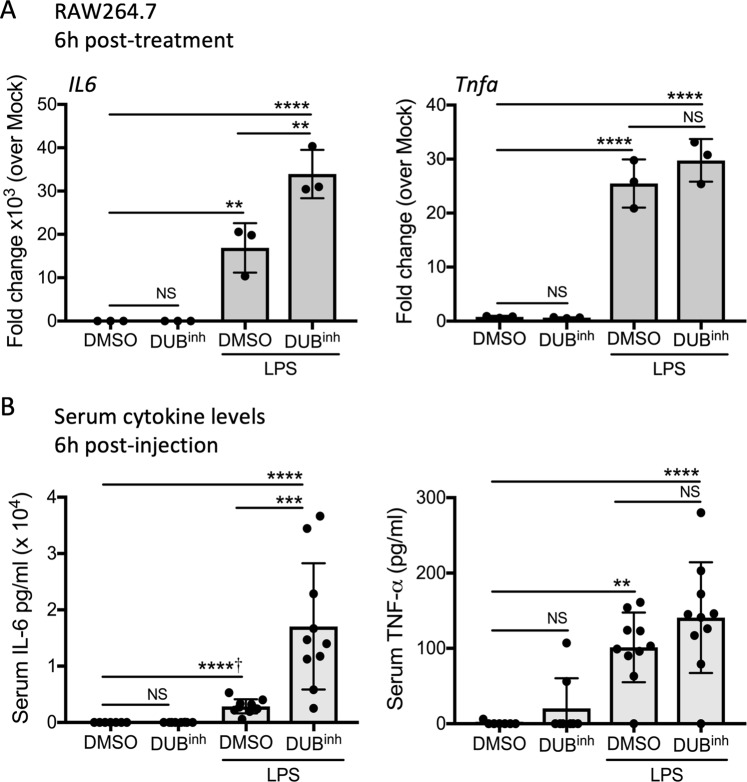
The innate immune system senses microbial ligands through pattern recognition and triggers downstream signaling cascades to promote inflammation and immune defense mechanisms. Emerging evidence suggests that cells also recognize alterations in host processes induced by infection as triggers. Protein ubiquitination and deubiquitination are post-translational modification processes essential for signaling and maintenance of cellular homeostasis, and infections can cause global alterations in the host ubiquitin proteome. Here we used a chemical biology approach to perturb the cellular ubiquitin proteome as a simplified model to study the impact of ubiquitin homeostasis alteration on macrophage function. Perturbation of ubiquitin homeostasis led to a rapid and transient burst of reactive oxygen species (ROS) that promoted macrophage inflammatory and anti-infective capacity. Moreover, we found that ROS production was dependent on the NOX2 phagocyte NADPH oxidase. Global alteration of the ubiquitin proteome also enhanced proinflammatory cytokine production in mice stimulated with a sub-lethal dose of LPS. Collectively, our findings suggest that major changes in the host ubiquitin landscape may be a potent signal to rapidly deploy innate immune defenses.
Conflict of interest statement
The authors declare no competing interests.
Figures
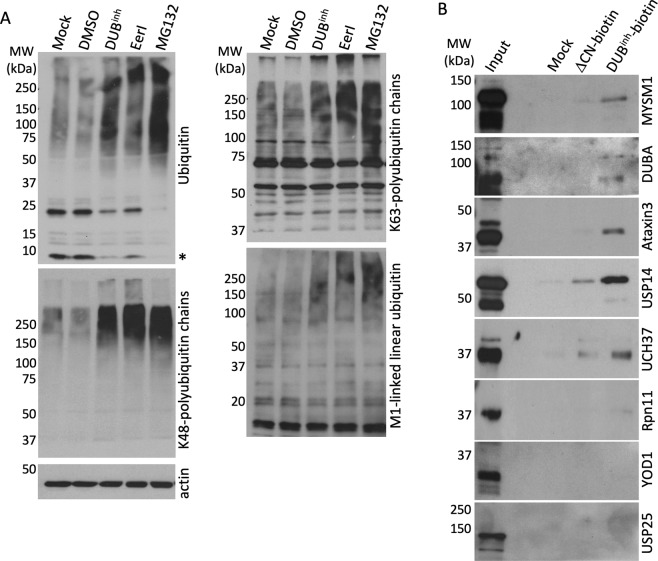
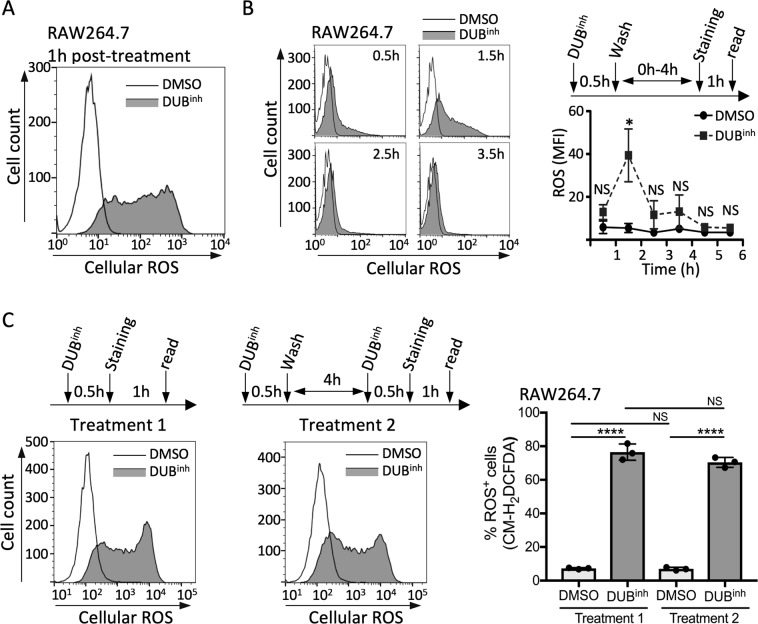
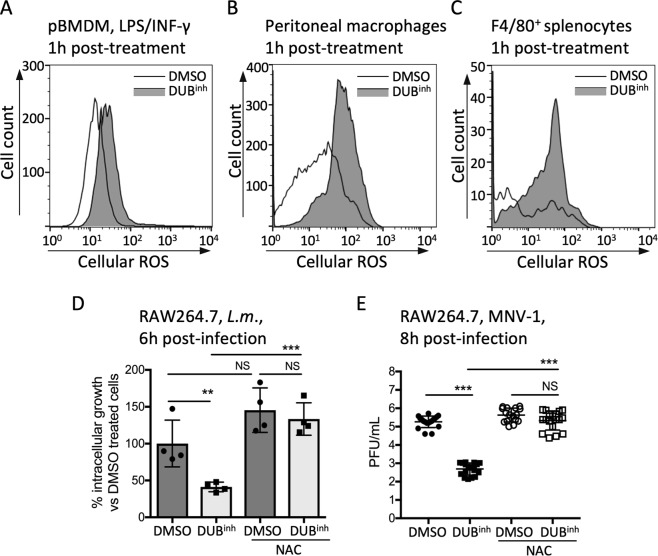
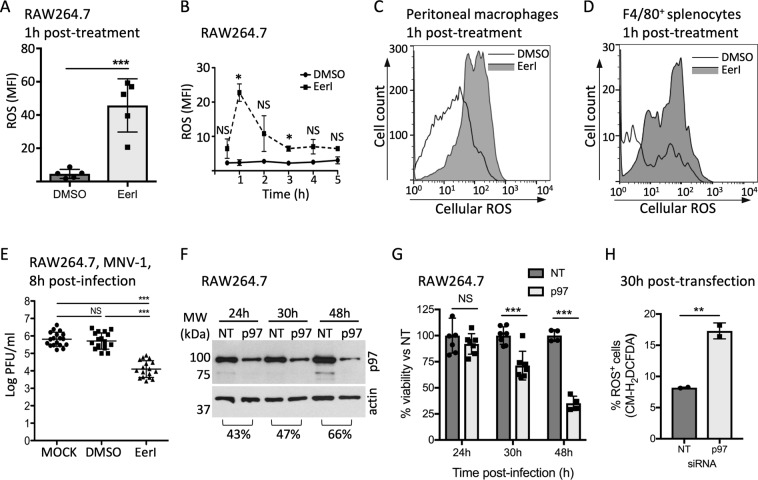
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
