

Evolutionary persistence of insect bunyavirus infection despite host acquisition and expression of the viral nucleoprotein gene
- PMID: 31308960
- PMCID: PMC6620529
- DOI: 10.1093/ve/vez017
Evolutionary persistence of insect bunyavirus infection despite host acquisition and expression of the viral nucleoprotein gene
Abstract
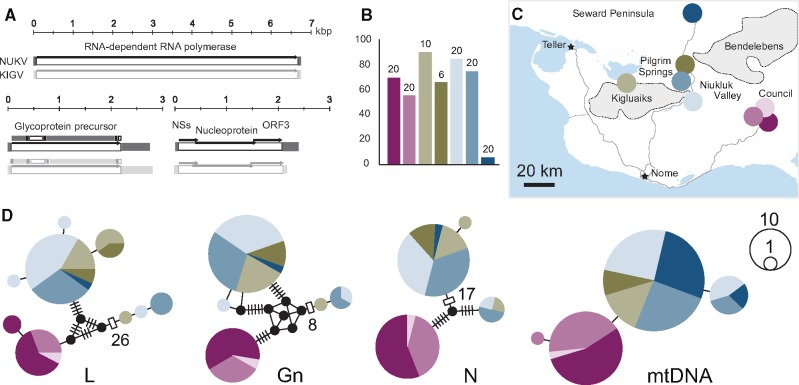
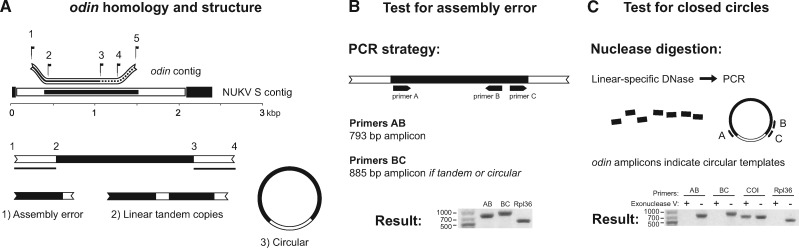
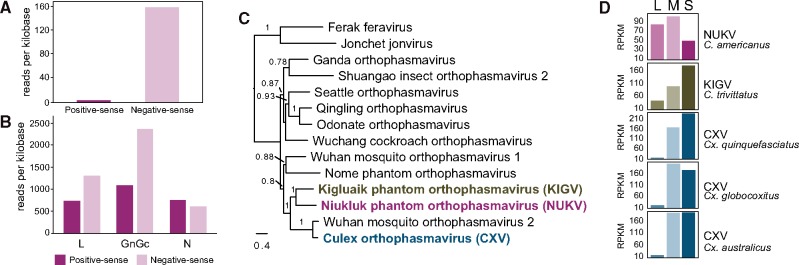
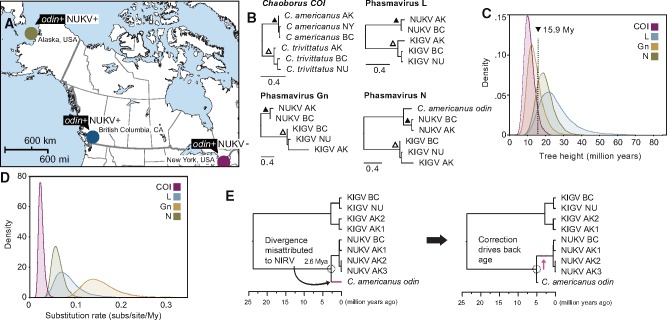
How insects combat RNA virus infection is a subject of intensive research owing to its importance in insect health, virus evolution, and disease transmission. In recent years, a pair of potentially linked phenomena have come to light as a result of this work-first, the pervasive production of viral DNA from exogenous nonretroviral RNA in infected individuals, and second, the widespread distribution of nonretroviral integrated RNA virus sequences (NIRVs) in the genomes of diverse eukaryotes. The evolutionary consequences of NIRVs for viruses are unclear and the field would benefit from studies of natural virus infections co-occurring with recent integrations, an exceedingly rare circumstance in the literature. Here, we provide evidence that a novel insect-infecting phasmavirus (Order Bunyavirales) has been persisting in a phantom midge host, Chaoborus americanus, for millions of years. Interestingly, the infection persists despite the host's acquisition (during the Pliocene), fixation, and expression of the viral nucleoprotein gene. We show that virus prevalence and geographic distribution are high and broad, comparable to the host-specific infections reported in other phantom midges. Short-read mapping analyses identified a lower abundance of the nucleoprotein-encoding genome segment in this virus relative to related viruses. Finally, the novel virus has facilitated the first substitution rate estimation for insect-infecting phasmaviruses. Over a period of approximately 16 million years, we find rates of (0.6 - 1.6) × 10-7 substitutions per site per year in protein coding genes, extraordinarily low for negative-sense RNA viruses, but consistent with the few estimates produced over comparable evolutionary timescales.
Keywords: EVEs; NIRVs; insect immunity; paleovirology; substitution rates.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
