

Mechanisms of Transcriptional Pausing in Bacteria
- PMID: 31310765
- PMCID: PMC6874753
- DOI: 10.1016/j.jmb.2019.07.017
Mechanisms of Transcriptional Pausing in Bacteria
Abstract
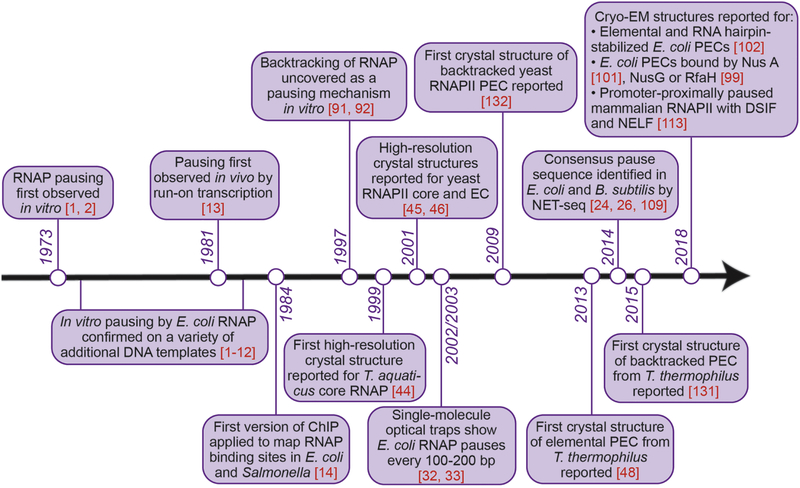
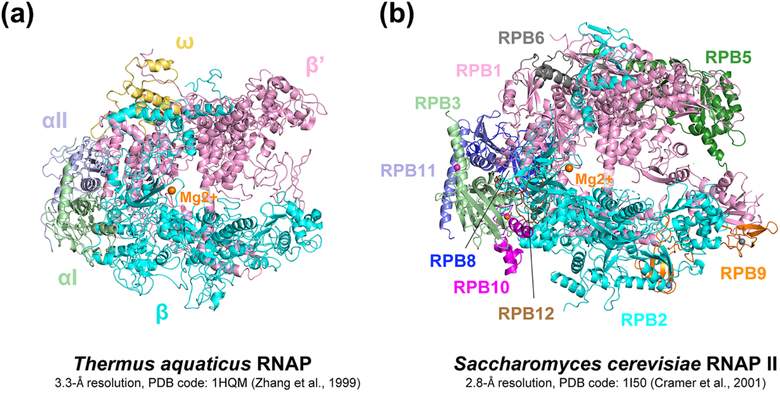
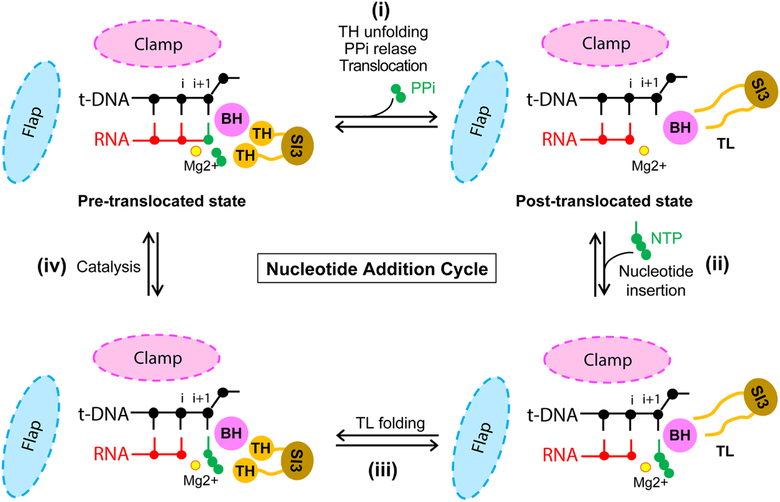
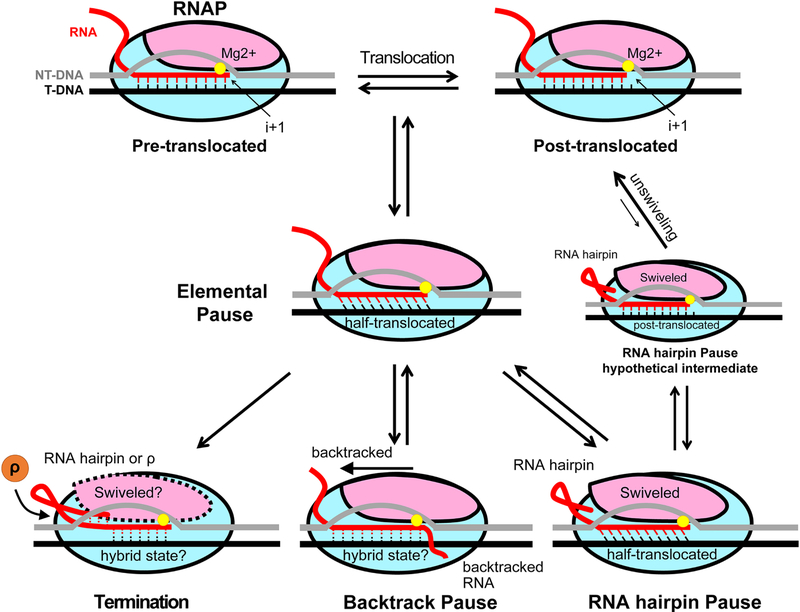
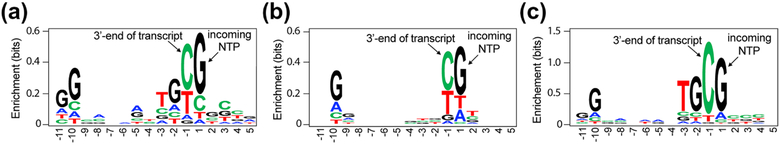
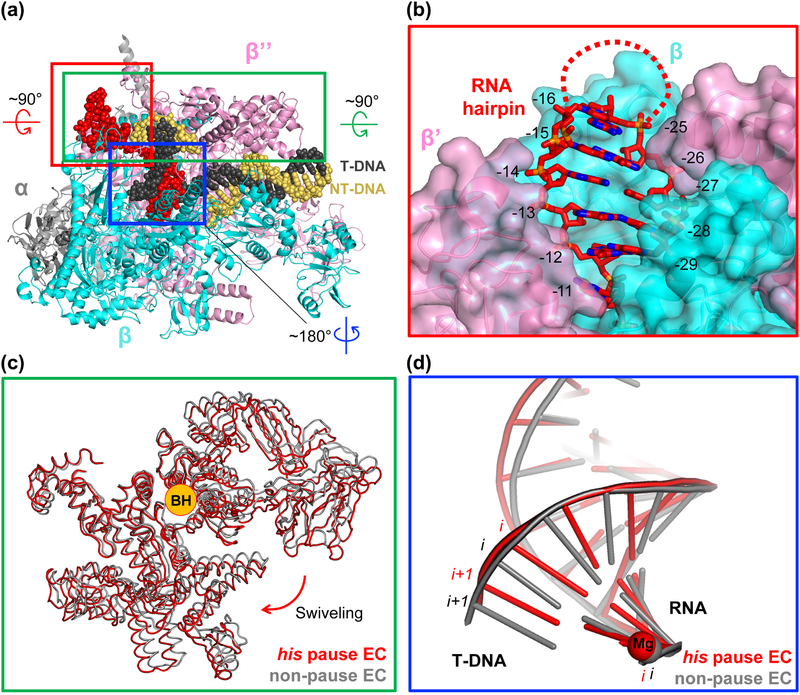
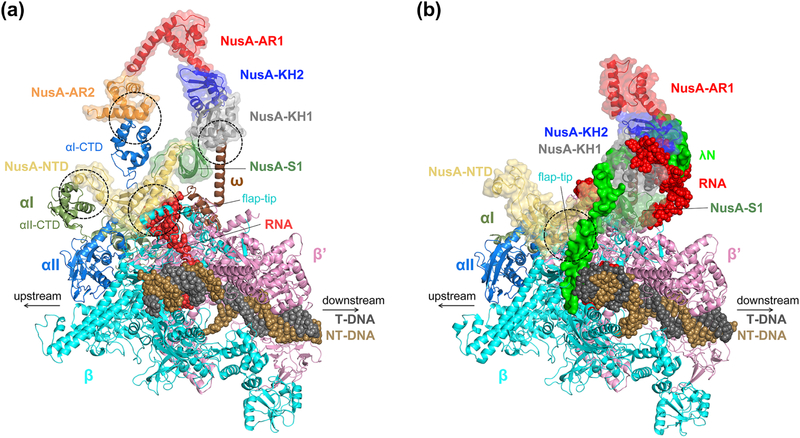
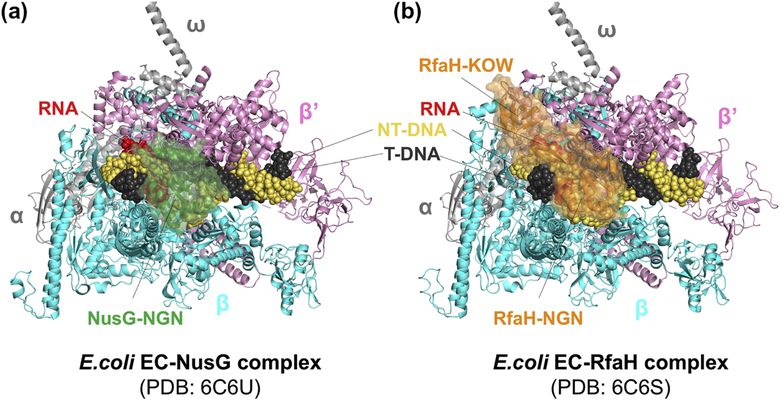
Pausing by RNA polymerase (RNAP) during transcription regulates gene expression in all domains of life. In this review, we recap the history of transcriptional pausing discovery, summarize advances in our understanding of the underlying causes of pausing since then, and describe new insights into the pausing mechanisms and pause modulation by transcription factors gained from structural and biochemical experiments. The accumulated evidence to date suggests that upon encountering a pause signal in the nucleic-acid sequence being transcribed, RNAP rearranges into an elemental, catalytically inactive conformer unable to load NTP substrate. The conformation, and as a consequence lifetime, of an elemental paused RNAP is modulated by backtracking, nascent RNA structure, binding of transcription regulators, or a combination of these mechanisms. We conclude the review by outlining open questions and directions for future research in the field of transcriptional pausing.
Keywords: Backtrack pause; Elemental pause; NusA; NusG; RNA hairpin pause.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
References
-
- Dahlberg JE, Blattner FR. Virus Res, ICN-UCLA Symp Mol Biol Proc, 2nd. 1973:533.
-
- Winkler ME, Yanofsky C. Pausing of RNA polymerase during in vitro transcription of the tryptophan operon leader region. Biochemistry. 1981;20:3738–44. - PubMed
-
- Lee F, Squires CL, Squires C, Yanofsky C. Termination of transcription in vitro in the escherichia coli tryptophan operon leader region. Journal of Molecular Biology. 1976;103:383–93. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
