

Cilantro leaf harbors a potent potassium channel-activating anticonvulsant
- PMID: 31311306
- PMCID: PMC6766653
- DOI: 10.1096/fj.201900485R
Cilantro leaf harbors a potent potassium channel-activating anticonvulsant
Abstract
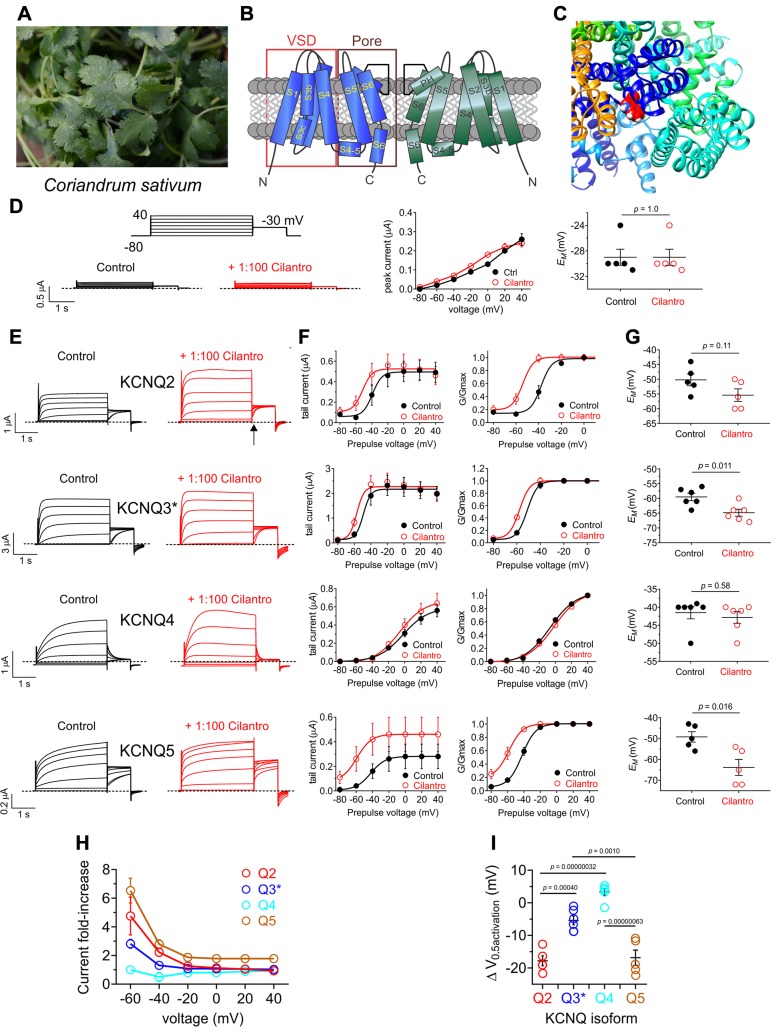
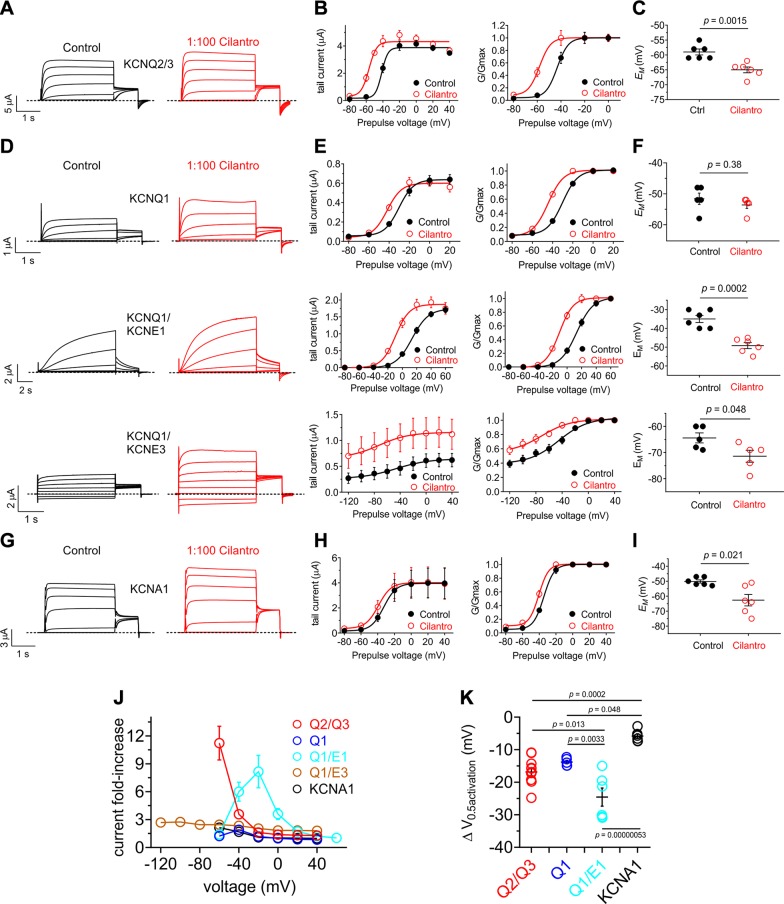
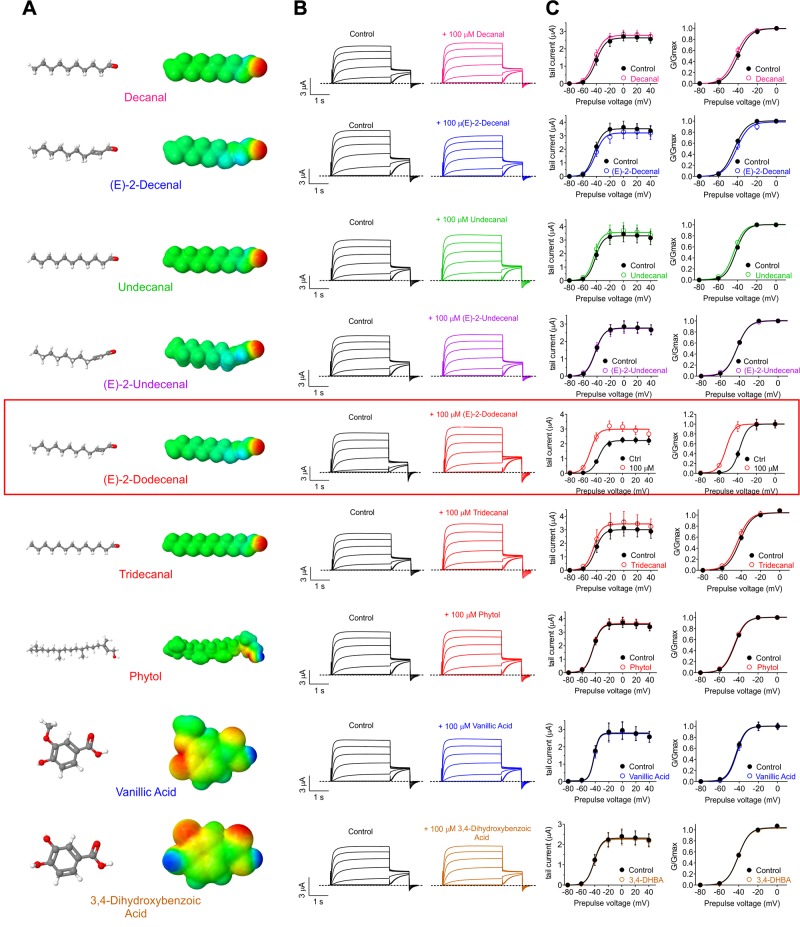
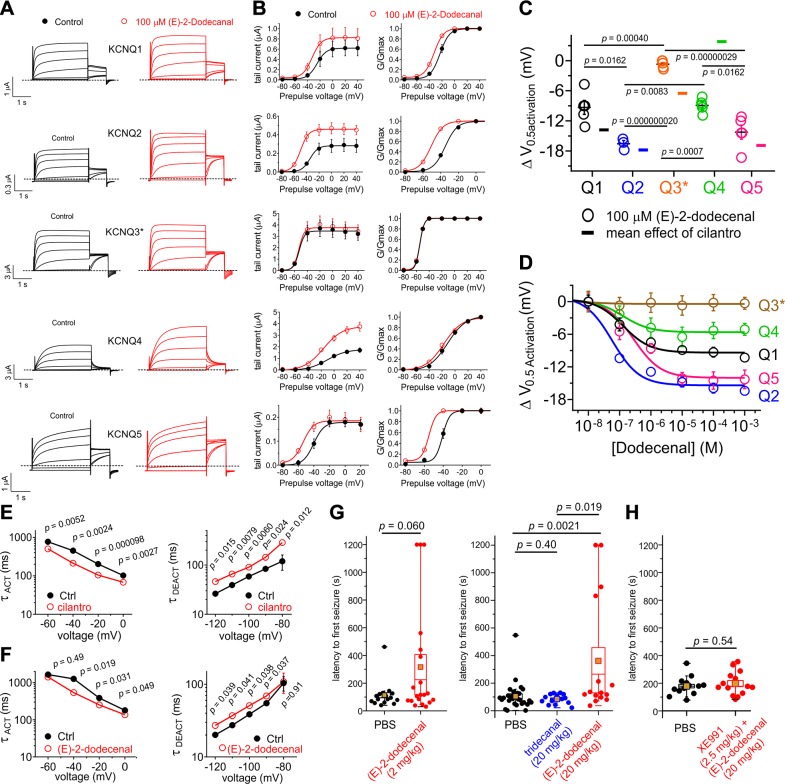
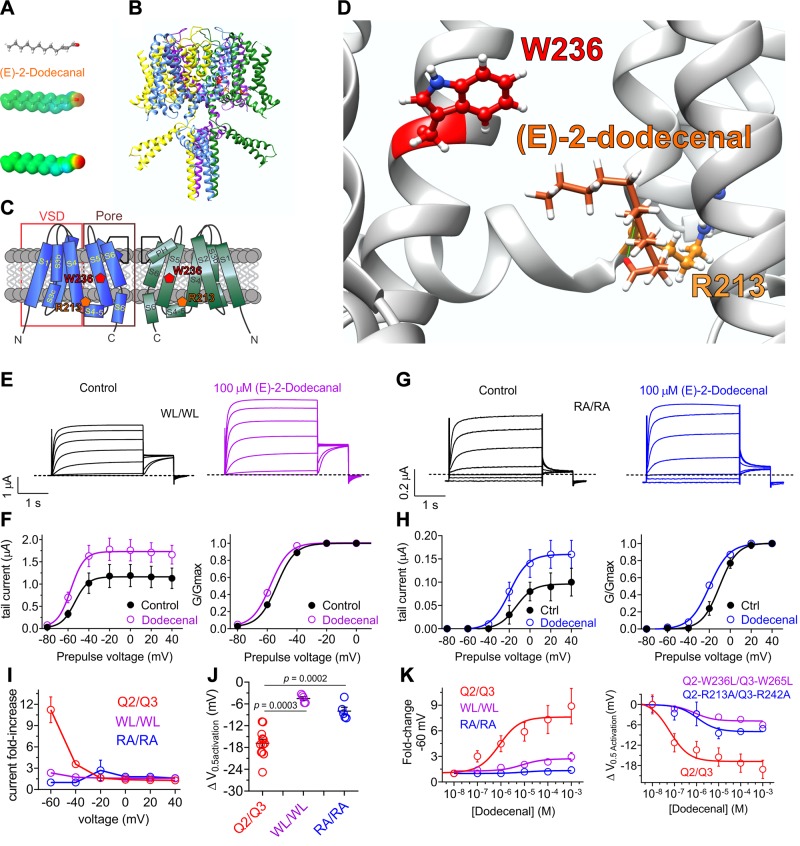
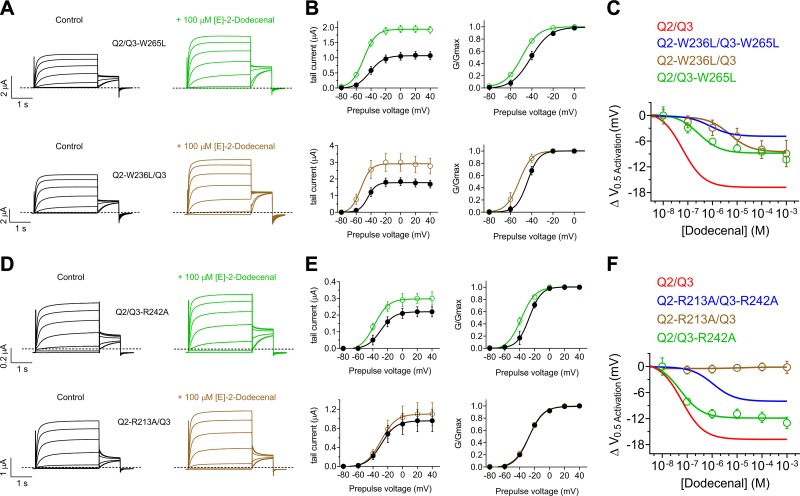
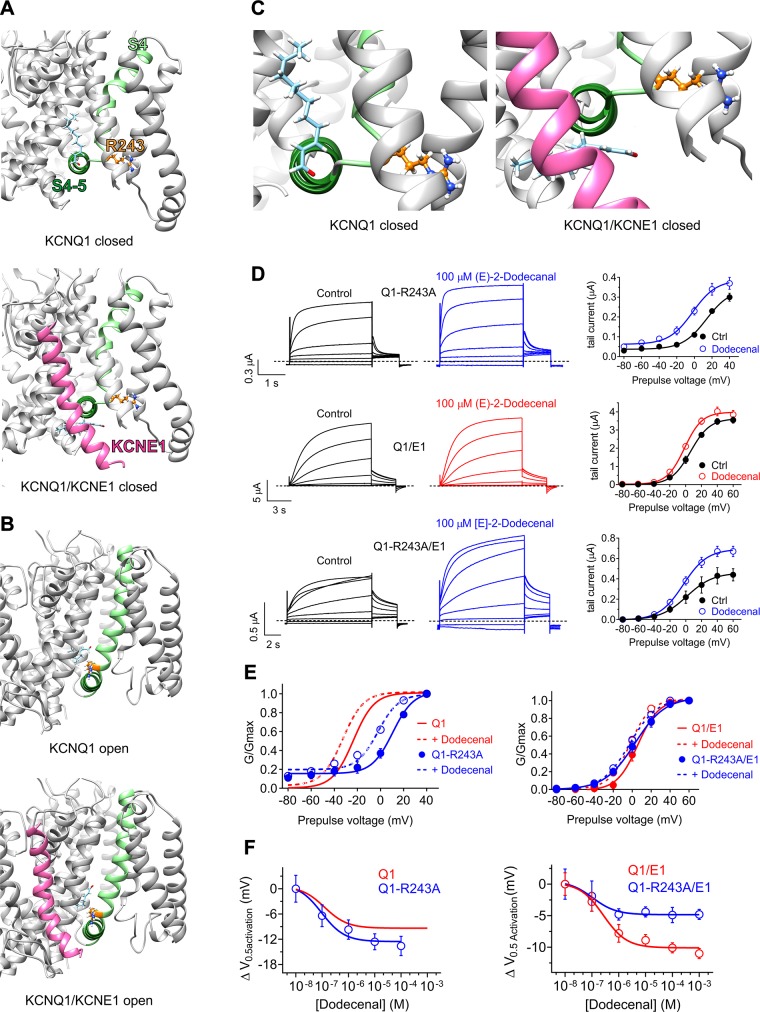
Herbs have a long history of use as folk medicine anticonvulsants, yet the underlying mechanisms often remain unknown. Neuronal voltage-gated potassium channel subfamily Q (KCNQ) dysfunction can cause severe epileptic encephalopathies that are resistant to modern anticonvulsants. Here we report that cilantro (Coriandrum sativum), a widely used culinary herb that also exhibits antiepileptic and other therapeutic activities, is a highly potent KCNQ channel activator. Screening of cilantro leaf metabolites revealed that one, the long-chain fatty aldehyde (E)-2-dodecenal, activates multiple KCNQs, including the predominant neuronal isoform, KCNQ2/KCNQ3 [half maximal effective concentration (EC50), 60 ± 20 nM], and the predominant cardiac isoform, KCNQ1 in complexes with the type I transmembrane ancillary subunit (KCNE1) (EC50, 260 ± 100 nM). (E)-2-dodecenal also recapitulated the anticonvulsant action of cilantro, delaying pentylene tetrazole-induced seizures. In silico docking and mutagenesis studies identified the (E)-2-dodecenal binding site, juxtaposed between residues on the KCNQ S5 transmembrane segment and S4-5 linker. The results provide a molecular basis for the therapeutic actions of cilantro and indicate that this ubiquitous culinary herb is surprisingly influential upon clinically important KCNQ channels.-Manville, R. W., Abbott, G. W. Cilantro leaf harbors a potent potassium channel-activating anticonvulsant.
Keywords: KCNQ1; KCNQ2; KCNQ3; epilepsy; herbal medicine.
Conflict of interest statement
The authors are grateful to Angele De Silva (University of California–Irvine) for generating mutant channel constructs, and to Bo Abbott for botanical photography. This study was supported by the U.S. National Institutes of Health (NIH) National Institute of General Medical Sciences (Grants GM115189 and GM130377 to G.W.A.) and NIH National Institute of Neurological Disorders and Stroke (Grant NS107671 to G.W.A.). The authors declare no conflicts of interest.
Figures
Similar articles
-
Ancient and modern anticonvulsants act synergistically in a KCNQ potassium channel binding pocket.Nat Commun. 2018 Sep 21;9(1):3845. doi: 10.1038/s41467-018-06339-2. Nat Commun. 2018. PMID: 30242262 Free PMC article.
-
Gabapentin Is a Potent Activator of KCNQ3 and KCNQ5 Potassium Channels.Mol Pharmacol. 2018 Oct;94(4):1155-1163. doi: 10.1124/mol.118.112953. Epub 2018 Jul 18. Mol Pharmacol. 2018. PMID: 30021858 Free PMC article.
-
Electrophysiological and pharmacological characterization of a novel and potent neuronal Kv7 channel opener SCR2682 for antiepilepsy.FASEB J. 2019 Aug;33(8):9154-9166. doi: 10.1096/fj.201802848RR. Epub 2019 May 7. FASEB J. 2019. PMID: 31063701
-
The mechanism of action of retigabine (ezogabine), a first-in-class K+ channel opener for the treatment of epilepsy.Epilepsia. 2012 Mar;53(3):412-24. doi: 10.1111/j.1528-1167.2011.03365.x. Epub 2012 Jan 5. Epilepsia. 2012. PMID: 22220513 Review.
-
The therapeutic potential of neuronal KCNQ channel modulators.Expert Opin Ther Targets. 2003 Dec;7(6):737-48. doi: 10.1517/14728222.7.6.737. Expert Opin Ther Targets. 2003. PMID: 14640909 Review.
Cited by
-
Discovery of a potent, Kv7.3-selective potassium channel opener from a Polynesian traditional botanical anticonvulsant.Commun Chem. 2024 Oct 10;7(1):233. doi: 10.1038/s42004-024-01318-9. Commun Chem. 2024. PMID: 39390220 Free PMC article.
-
The GluN2B-Trp373 NMDA Receptor Variant is Associated with Autism-, Epilepsy-Related Phenotypes and Reduces NMDA Receptor Currents in Rats.Neurochem Res. 2022 Jun;47(6):1588-1597. doi: 10.1007/s11064-022-03554-8. Epub 2022 Feb 18. Neurochem Res. 2022. PMID: 35181828
-
Coriandrum sativum and Its Utility in Psychiatric Disorders.Molecules. 2023 Jul 10;28(14):5314. doi: 10.3390/molecules28145314. Molecules. 2023. PMID: 37513187 Free PMC article. Review.
-
Ancient medicinal plant rosemary contains a highly efficacious and isoform-selective KCNQ potassium channel opener.Commun Biol. 2023 Jun 15;6(1):644. doi: 10.1038/s42003-023-05021-8. Commun Biol. 2023. PMID: 37322081 Free PMC article.
-
KCNQ5 Potassium Channel Activation Underlies Vasodilation by Tea.Cell Physiol Biochem. 2021 Mar 6;55(S3):46-64. doi: 10.33594/000000337. Cell Physiol Biochem. 2021. PMID: 33667331 Free PMC article.
References
-
- Inskeep R. R. (1969) Health hazards and healing in antiquity. S. Afr. Archaeol. Bull. 24, 21–29
-
- Hardy K., Buckley S., Collins M. J., Estalrrich A., Brothwell D., Copeland L., García-Tabernero A., García-Vargas S., de la Rasilla M., Lalueza-Fox C., Huguet R., Bastir M., Santamaría D., Madella M., Wilson J., Cortés A. F., Rosas A. (2012) Neanderthal medics? Evidence for food, cooking, and medicinal plants entrapped in dental calculus. Naturwissenschaften 99, 617–626 - PubMed
-
- Weyrich L. S., Duchene S., Soubrier J., Arriola L., Llamas B., Breen J., Morris A. G., Alt K. W., Caramelli D., Dresely V., Farrell M., Farrer A. G., Francken M., Gully N., Haak W., Hardy K., Harvati K., Held P., Holmes E. C., Kaidonis J., Lalueza-Fox C., de la Rasilla M., Rosas A., Semal P., Soltysiak A., Townsend G., Usai D., Wahl J., Huson D. H., Dobney K., Cooper A. (2017) Neanderthal behaviour, diet, and disease inferred from ancient DNA in dental calculus. Nature 544, 357–361 - PubMed
-
- Hoffmann D. (2003) Medical Herbalism: The Science and Practice of Herbal Medicine, Healing Arts Press, Rochester, VT, USA
