

miR-221 and -222 target CACNA1C and KCNJ5 leading to altered cardiac ion channel expression and current density
- PMID: 31312877
- PMCID: PMC7058603
- DOI: 10.1007/s00018-019-03217-y
miR-221 and -222 target CACNA1C and KCNJ5 leading to altered cardiac ion channel expression and current density
Abstract
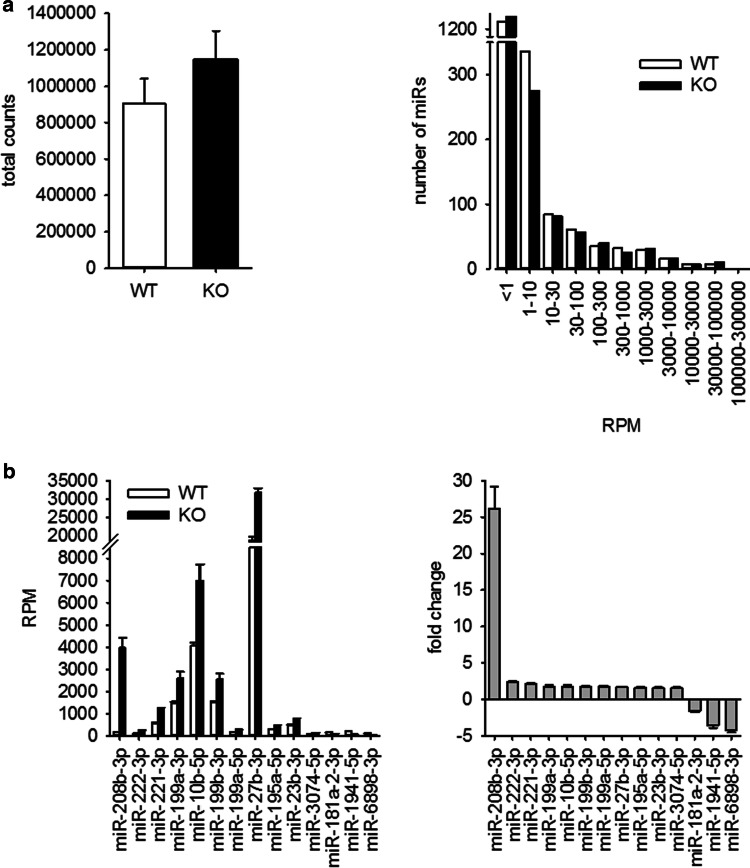
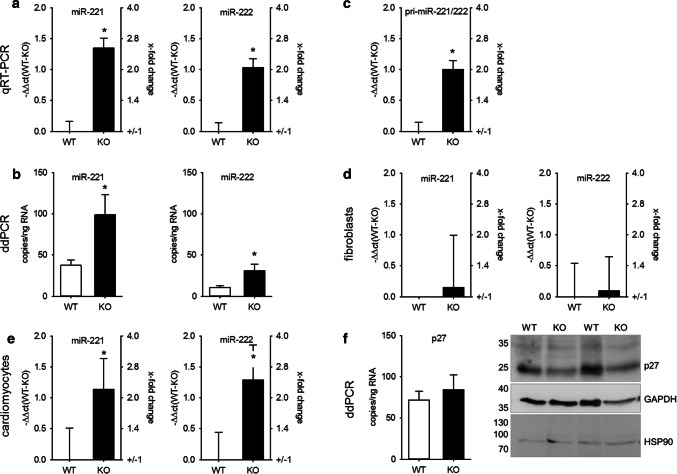
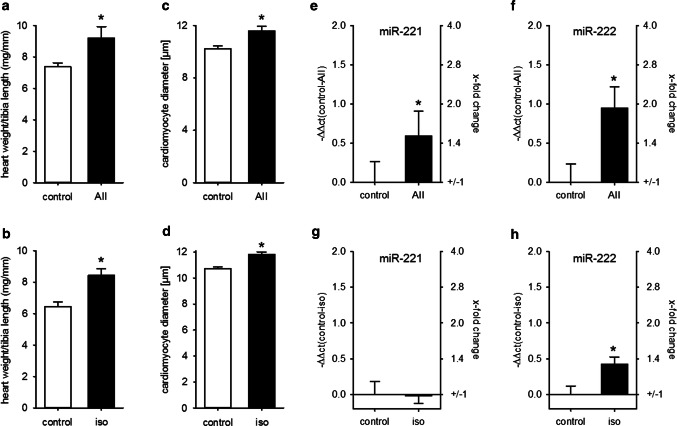
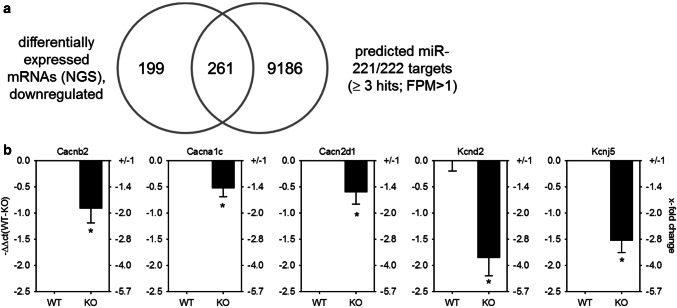
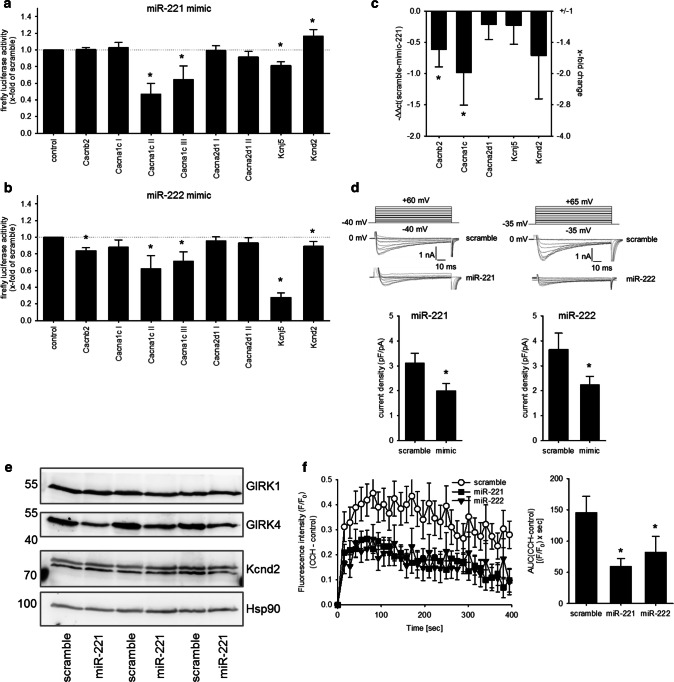
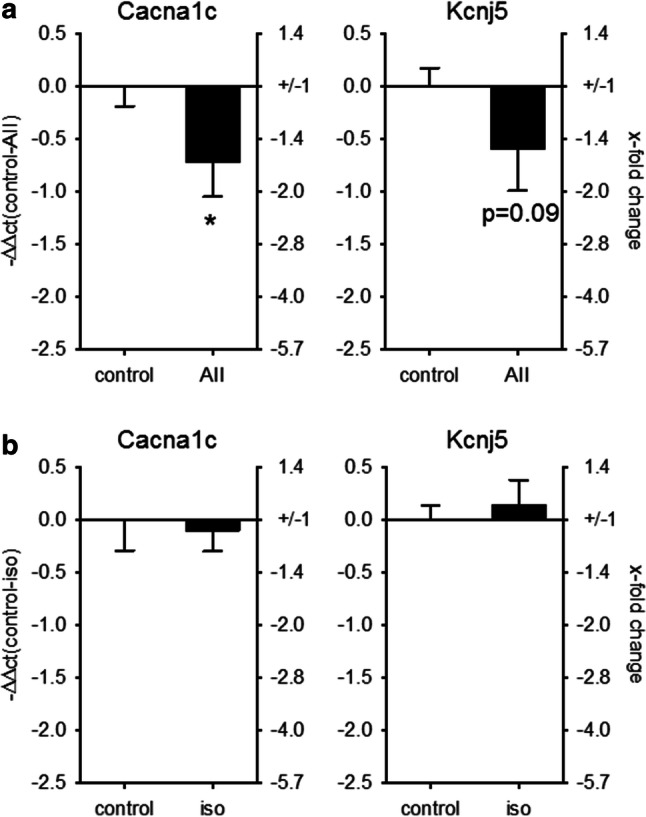
MicroRNAs (miRs) contribute to different aspects of cardiovascular pathology, among others cardiac hypertrophy and atrial fibrillation. The aim of our study was to evaluate the impact of miR-221/222 on cardiac electrical remodeling. Cardiac miR expression was analyzed in a mouse model with altered electrocardiography parameters and severe heart hypertrophy. Next generation sequencing revealed 14 differentially expressed miRs in hypertrophic hearts, with miR-221 and -222 being the strongest regulated miR-cluster. This increase was restricted to cardiomyocytes and not observed in cardiac fibroblasts. Additionally, we evaluated the change of miR-221/222 in vivo in two models of pharmacologically induced heart hypertrophy (angiotensin II, isoprenaline), thereby demonstrating a stimulus-induced increase in miR-221/222 in vivo by angiotensin II but not by isoprenaline. Whole transcriptome analysis by RNA-seq and qRT-PCR validation revealed an enriched number of downregulated mRNAs coding for proteins located in the T-tubule, which are also predicted targets for miR-221/222. Among those, mRNAs were the L-type Ca2+ channel subunits as well as potassium channel subunits. We confirmed that both miRs target the 3'-untranslated regions of Cacna1c and Kcnj5. Furthermore, enhanced expression of these miRs reduced L-type Ca2+ channel and Kcnj5 channel abundance and function, which was analyzed by whole-cell patch clamp recordings or Western blot and flux measurements, respectively. miR-221 and -222 contribute to the regulation of L-type Ca2+ channels as well as Kcnj5 channels and, therefore, potentially contribute to disturbed cardiac excitation generation and propagation. Future studies will have to evaluate the pathophysiological and clinical relevance of aberrant miR-221/222 expression for electrical remodeling.
Keywords: Angiotensin II; Cardiomyocytes; Electrical remodeling; Heart hypertrophy.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
