

Prefrontal Pathways Provide Top-Down Control of Memory for Sequences of Events
- PMID: 31315044
- PMCID: PMC6662648
- DOI: 10.1016/j.celrep.2019.06.053
Prefrontal Pathways Provide Top-Down Control of Memory for Sequences of Events
Abstract
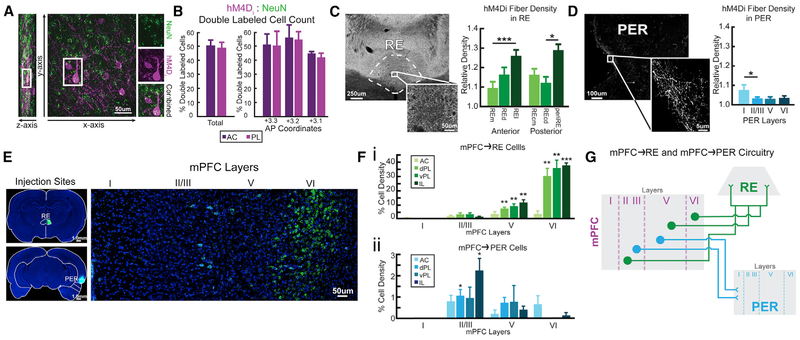
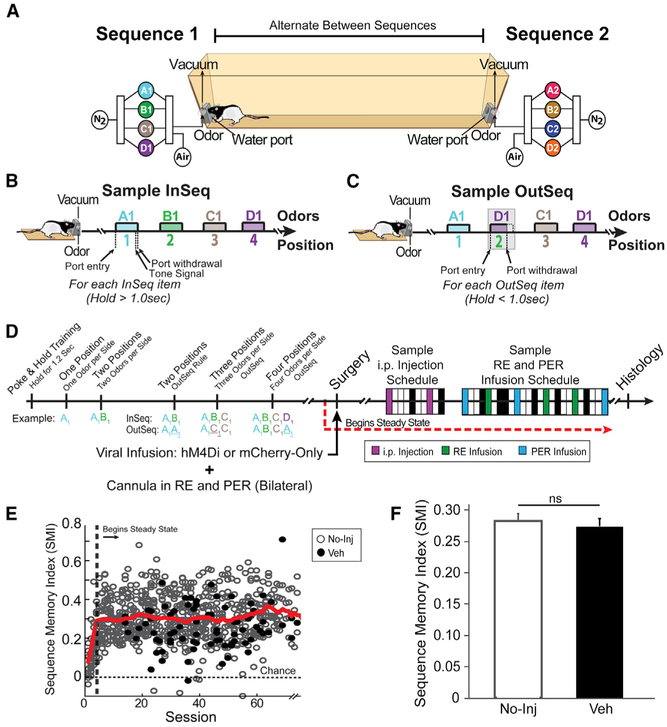
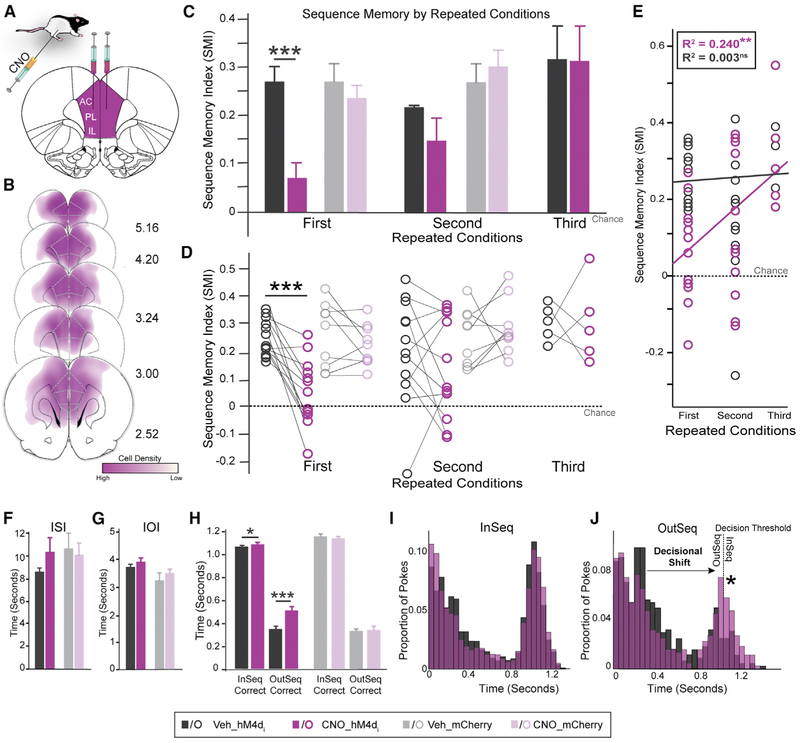
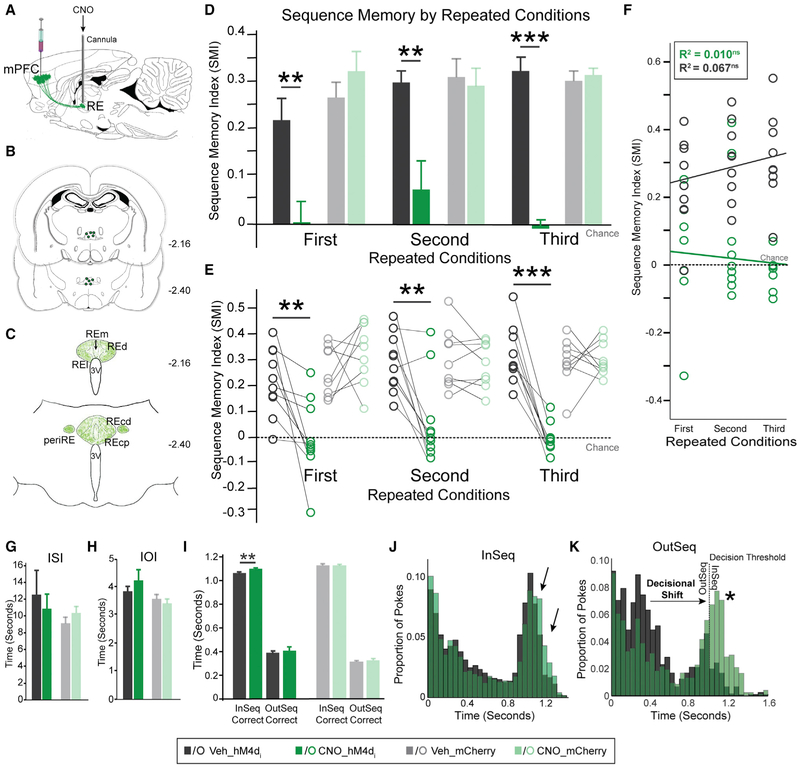
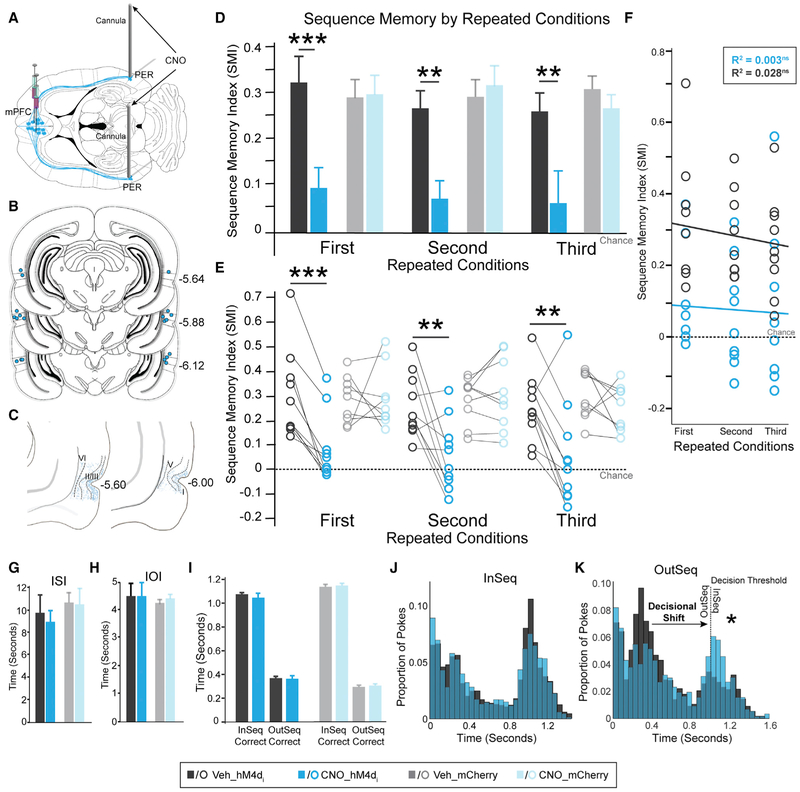
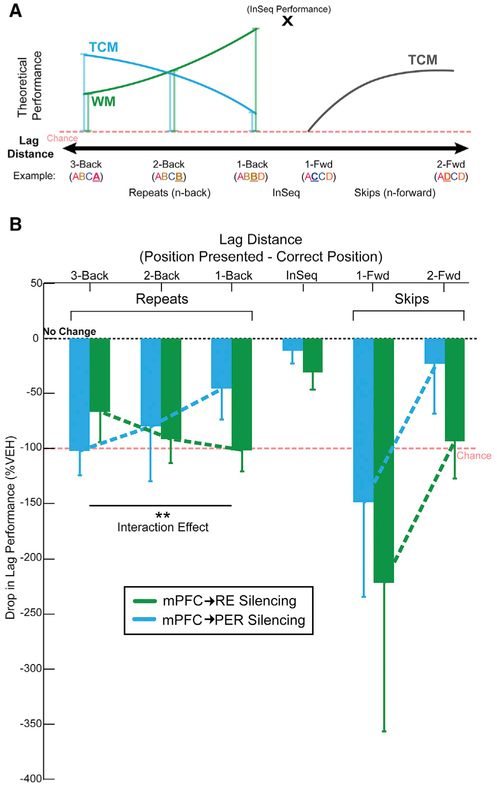
We remember our lives as sequences of events, but it is unclear how these memories are controlled during retrieval. In rats, the medial prefrontal cortex (mPFC) is positioned to influence sequence memory through extensive top-down inputs to regions heavily interconnected with the hippocampus, notably the nucleus reuniens of the thalamus (RE) and perirhinal cortex (PER). Here, we used an hM4Di synaptic-silencing approach to test our hypothesis that specific mPFC→RE and mPFC→PER projections regulate sequence memory retrieval. First, we found non-overlapping populations of mPFC cells project to RE and PER. Second, suppressing mPFC activity impaired sequence memory. Third, inhibiting mPFC→RE and mPFC→PER pathways effectively abolished sequence memory. Finally, a sequential lag analysis showed that the mPFC→RE pathway contributes to a working memory retrieval strategy, whereas the mPFC→PER pathway supports a temporal context memory retrieval strategy. These findings demonstrate that mPFC→RE and mPFC→PER pathways serve as top-down mechanisms that control distinct sequence memory retrieval strategies.
Keywords: DREADDs; cognitive control; episodic memory; hippocampus; memory retrieval; nucleus reuniens; perirhinal cortex; temporal context; thalamus; working memory.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
