

Human ATG4 autophagy proteases counteract attachment of ubiquitin-like LC3/GABARAP proteins to other cellular proteins
- PMID: 31315929
- PMCID: PMC6709618
- DOI: 10.1074/jbc.AC119.009977
Human ATG4 autophagy proteases counteract attachment of ubiquitin-like LC3/GABARAP proteins to other cellular proteins
Abstract
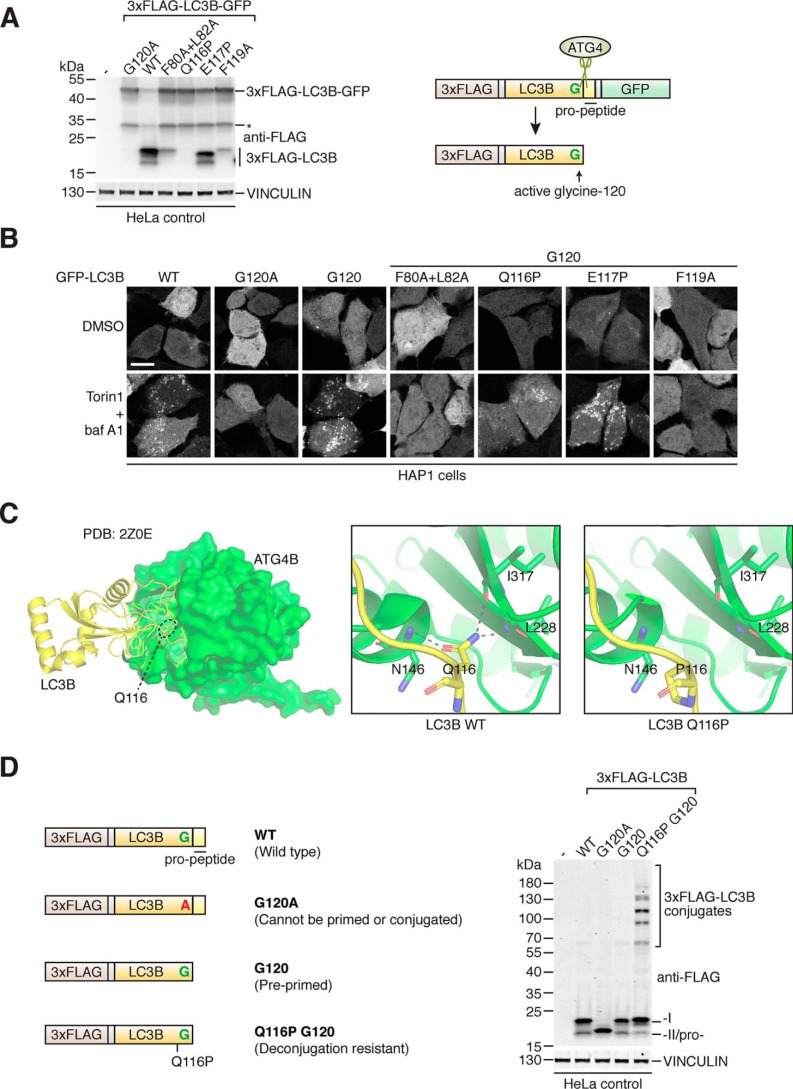
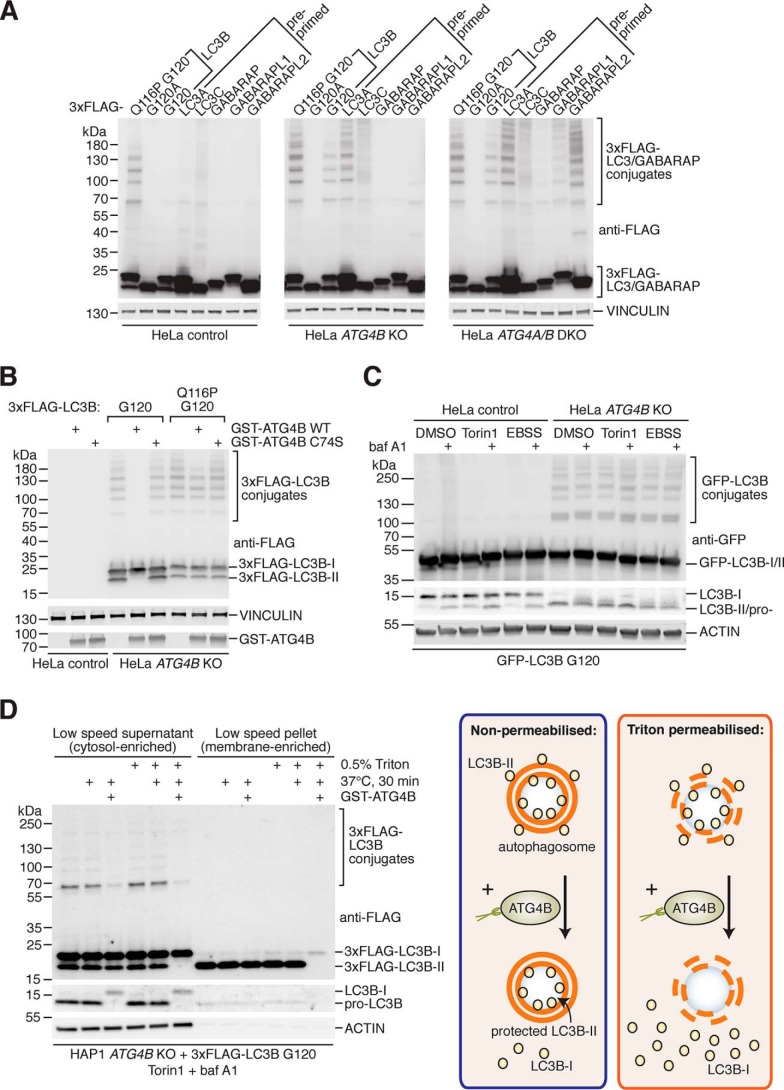
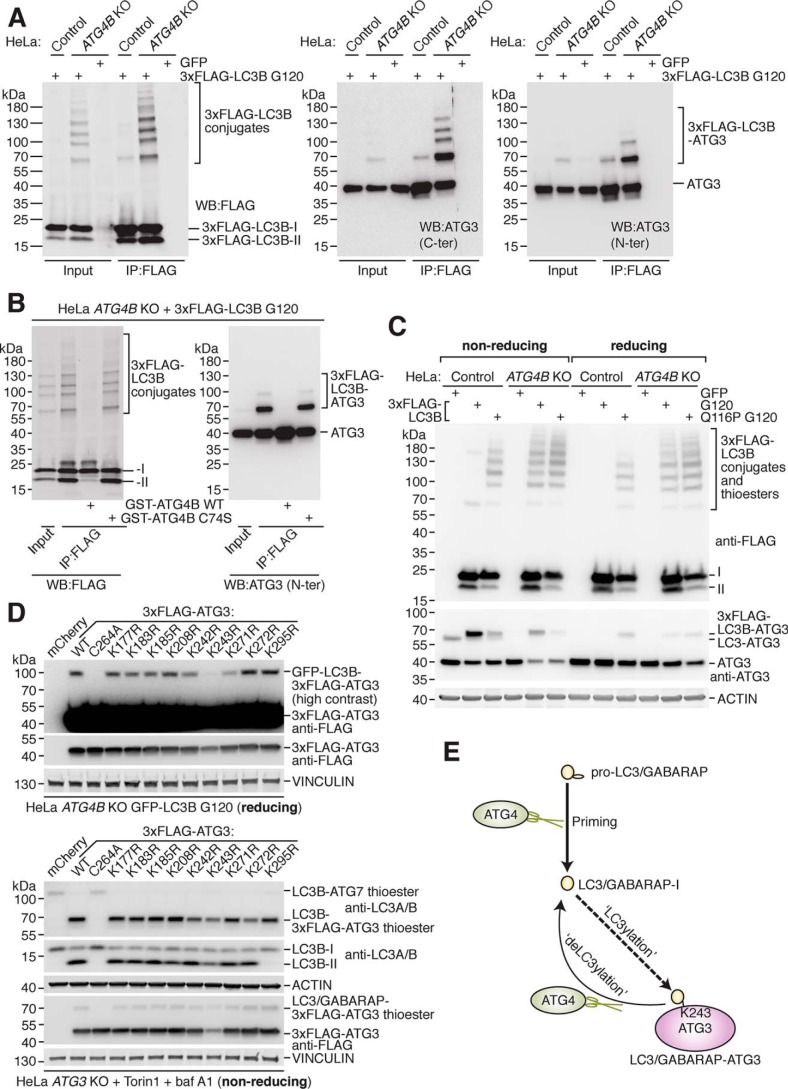
Microtubule-associated protein 1 light chain 3 α (LC3)/GABA type A receptor-associated protein (GABARAP) comprises a family of ubiquitin-like proteins involved in (macro)autophagy, an important intracellular degradation pathway that delivers cytoplasmic material to lysosomes via double-membrane vesicles called autophagosomes. The only currently known cellular molecules covalently modified by LC3/GABARAP are membrane phospholipids such as phosphatidylethanolamine in the autophagosome membrane. Autophagy-related 4 cysteine peptidase (ATG4) proteases process inactive pro-LC3/GABARAP before lipidation, and the same proteases can also deconjugate LC3/GABARAP from lipids. To determine whether LC3/GABARAP has other molecular targets, here we generated a pre-processed LC3B mutant (Q116P) that is resistant to ATG4-mediated deconjugation. Upon expression in human cells and when assessed by immunoblotting under reducing and denaturing conditions, deconjugation-resistant LC3B accumulated in multiple forms and at much higher molecular weights than free LC3B. We observed a similar accumulation when pre-processed versions of all mammalian LC3/GABARAP isoforms were expressed in ATG4-deficient cell lines, suggesting that LC3/GABARAP can attach also to other larger molecules. We identified ATG3, the E2-like enzyme involved in LC3/GABARAP lipidation, as one target of conjugation with multiple copies of LC3/GABARAP. We show that LC3B-ATG3 conjugates are distinct from the LC3B-ATG3 thioester intermediate formed before lipidation, and we biochemically demonstrate that ATG4B can cleave LC3B-ATG3 conjugates. Finally, we determined ATG3 residue Lys-243 as an LC3B modification site. Overall, we provide the first cellular evidence that mammalian LC3/GABARAP post-translationally modifies proteins akin to ubiquitination ("LC3ylation"), with ATG4 proteases acting like deubiquitinating enzymes to counteract this modification ("deLC3ylation").
Keywords: ATG4B; Atg8; GABARAPL2; LC3ylation; autophagy; cysteine protease; deconjugation; deubiquitylation (deubiquitination); post-translational modification; ubiquitin-conjugating enzyme (E2 enzyme).
© 2019 Agrotis et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Galluzzi L., Baehrecke E. H., Ballabio A., Boya P., Bravo-San Pedro J. M., Cecconi F., Choi A. M., Chu C. T., Codogno P., Colombo M. I., Cuervo A. M., Debnath J., Deretic V., Dikic I., Eskelinen E.-L., et al. (2017) Molecular definitions of autophagy and related processes. EMBO J. 36, 1811–1836 10.15252/embj.201796697 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
