

A live-attenuated RhCMV/SIV vaccine shows long-term efficacy against heterologous SIV challenge
- PMID: 31316007
- PMCID: PMC6788755
- DOI: 10.1126/scitranslmed.aaw2607
A live-attenuated RhCMV/SIV vaccine shows long-term efficacy against heterologous SIV challenge
Abstract
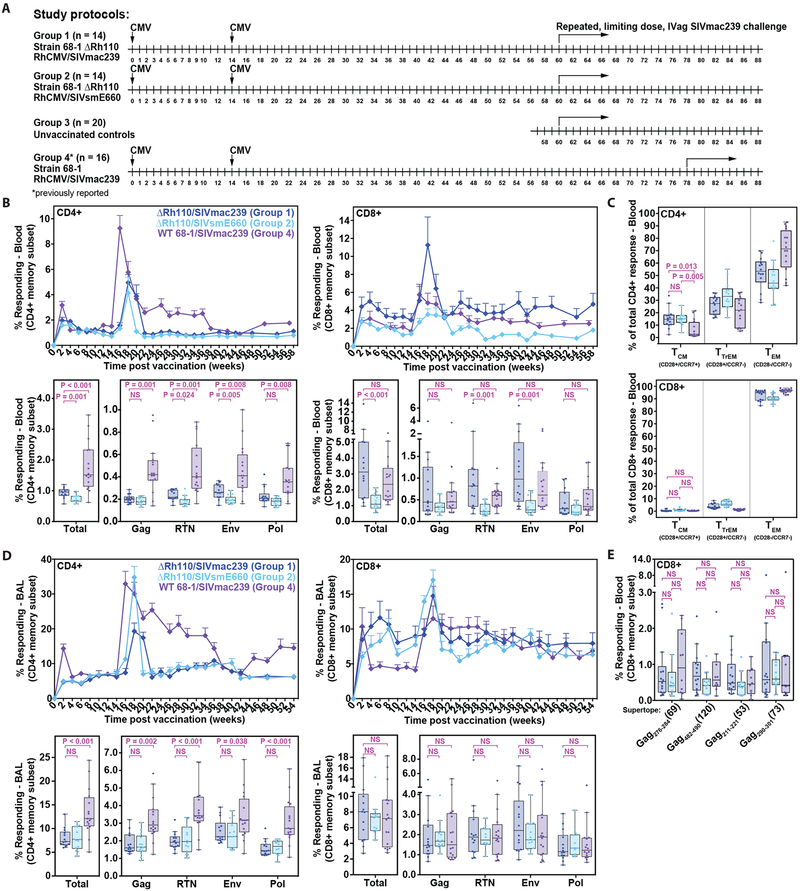
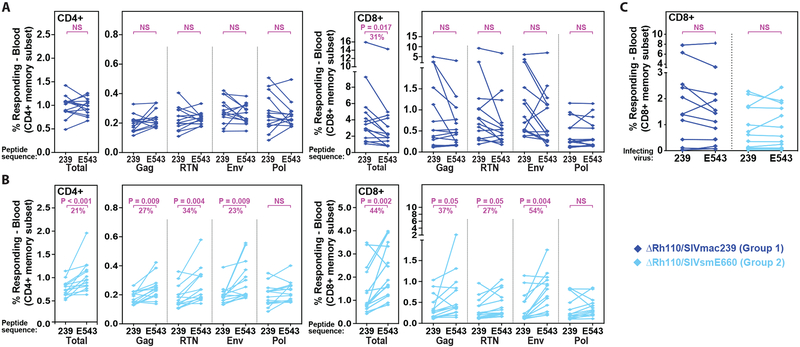
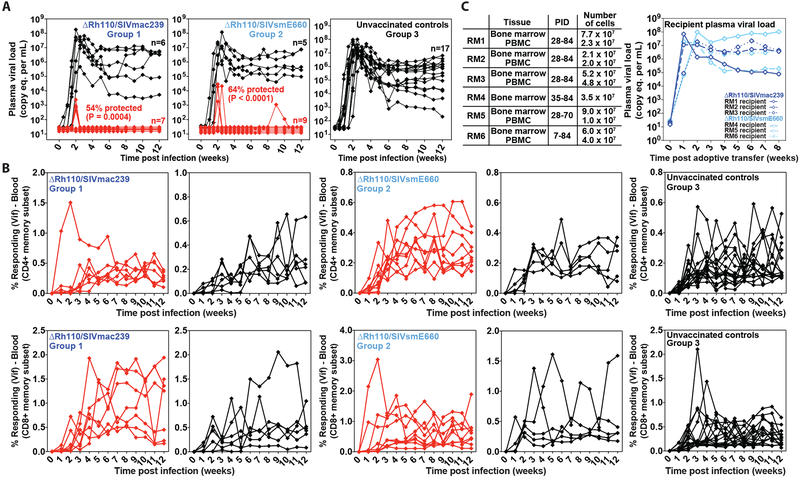
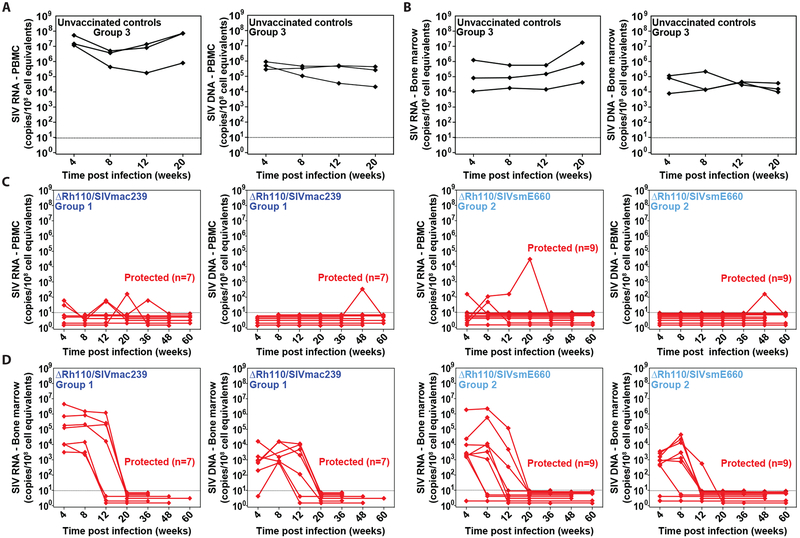
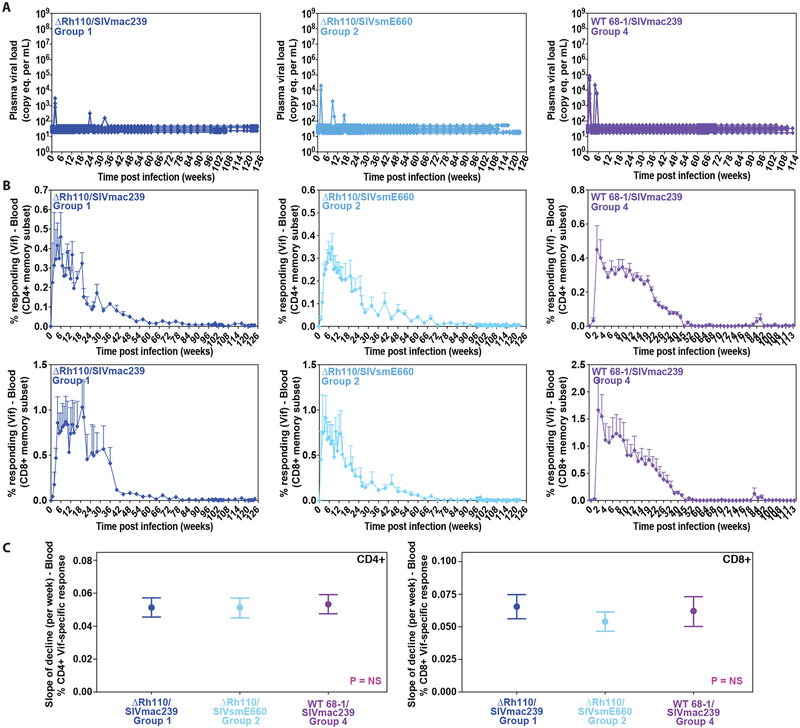
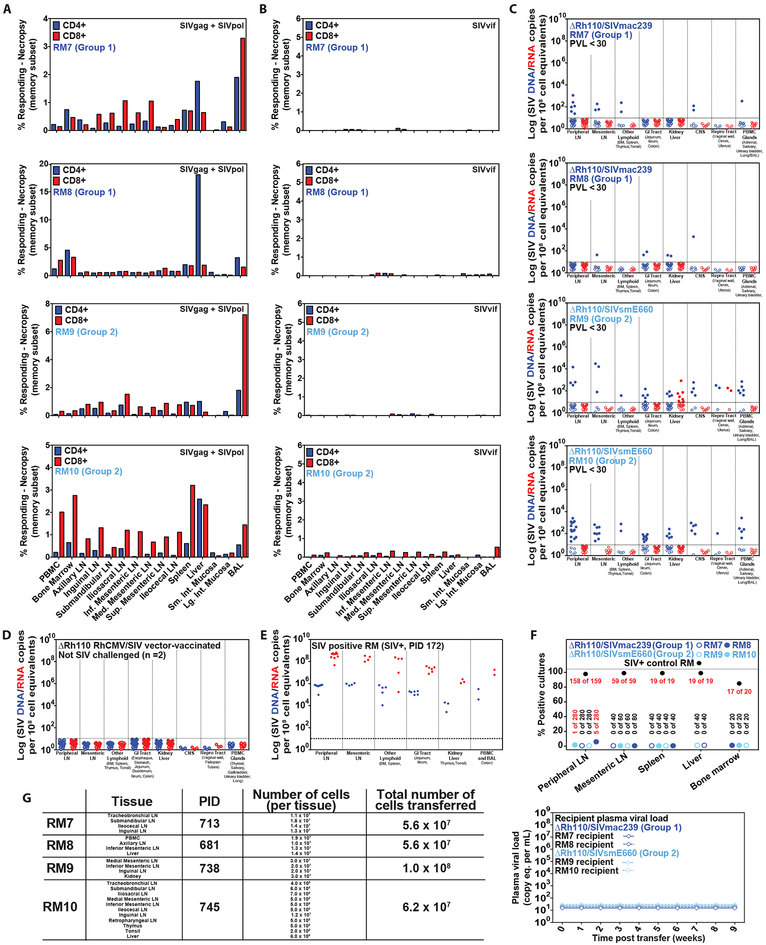
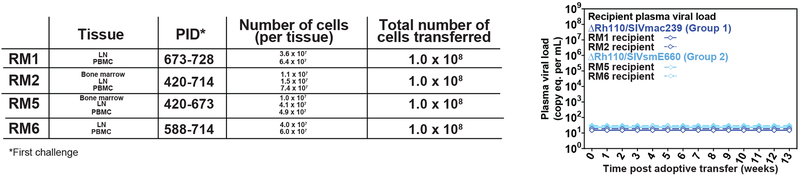
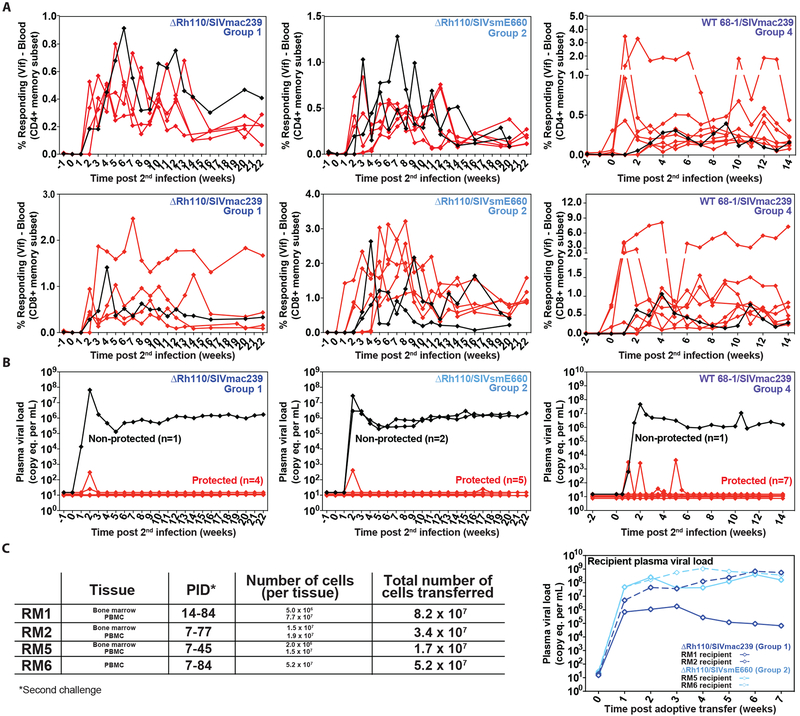
Previous studies have established that strain 68-1-derived rhesus cytomegalovirus (RhCMV) vectors expressing simian immunodeficiency virus (SIV) proteins (RhCMV/SIV) are able to elicit and maintain cellular immune responses that provide protection against mucosal challenge of highly pathogenic SIV in rhesus monkeys (RMs). However, these efficacious RhCMV/SIV vectors were replication and spread competent and therefore have the potential to cause disease in immunocompromised subjects. To develop a safer CMV-based vaccine for clinical use, we attenuated 68-1 RhCMV/SIV vectors by deletion of the Rh110 gene encoding the pp71 tegument protein (ΔRh110), allowing for suppression of lytic gene expression. ΔRh110 RhCMV/SIV vectors are highly spread deficient in vivo (~1000-fold compared to the parent vector) yet are still able to superinfect RhCMV+ RMs and generate high-frequency effector-memory-biased T cell responses. Here, we demonstrate that ΔRh110 68-1 RhCMV/SIV-expressing homologous or heterologous SIV antigens are highly efficacious against intravaginal (IVag) SIVmac239 challenge, providing control and progressive clearance of SIV infection in 59% of vaccinated RMs. Moreover, among 12 ΔRh110 RhCMV/SIV-vaccinated RMs that controlled and progressively cleared an initial SIV challenge, 9 were able to stringently control a second SIV challenge ~3 years after last vaccination, demonstrating the durability of this vaccine. Thus, ΔRh110 RhCMV/SIV vectors have a safety and efficacy profile that warrants adaptation and clinical evaluation of corresponding HCMV vectors as a prophylactic HIV/AIDS vaccine.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Stover J, Hallett TB, Wu Z, Warren M, Gopalappa C, Pretorius C, Ghys PD, Montaner J, Schwartlander B, New G Prevention Technology Study, How can we get close to zero? The potential contribution of biomedical prevention and the investment framework towards an effective response to HIV. PLoS One 9, e111956 (2014). - PMC - PubMed
-
- Jarvis MA, Hansen SG, Nelson JA, Picker LJ, Früh K, in Cytomegaloviruses: From Molecular Pathogenesis to Intervention Reddehase MJ, Ed. (Caister Academic Press, 2013), vol. 2, chap. 21.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
