

Stem cells in tissues, organoids, and cancers
- PMID: 31317205
- PMCID: PMC6785598
- DOI: 10.1007/s00018-019-03199-x
Stem cells in tissues, organoids, and cancers
Abstract
Stem cells give rise to all cells and build the tissue structures in our body, and heterogeneity and plasticity are the hallmarks of stem cells. Epigenetic modification, which is associated with niche signals, determines stem cell differentiation and somatic cell reprogramming. Stem cells play a critical role in the development of tumors and are capable of generating 3D organoids. Understanding the properties of stem cells will improve our capacity to maintain tissue homeostasis. Dissecting epigenetic regulation could be helpful for achieving efficient cell reprograming and for developing new drugs for cancer treatment. Stem cell-derived organoids open up new avenues for modeling human diseases and for regenerative medicine. Nevertheless, in addition to the achievements in stem cell research, many challenges still need to be overcome for stem cells to have versatile application in clinics.
Keywords: Cancer stem cell; Organoid; Stem cell; Stem cell niche.
Figures
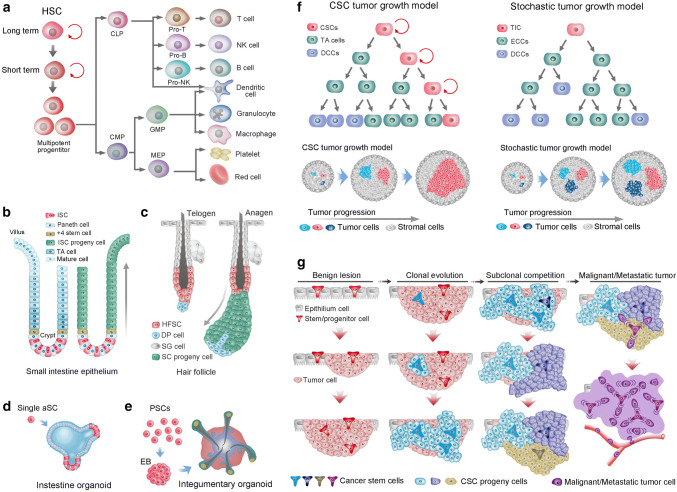
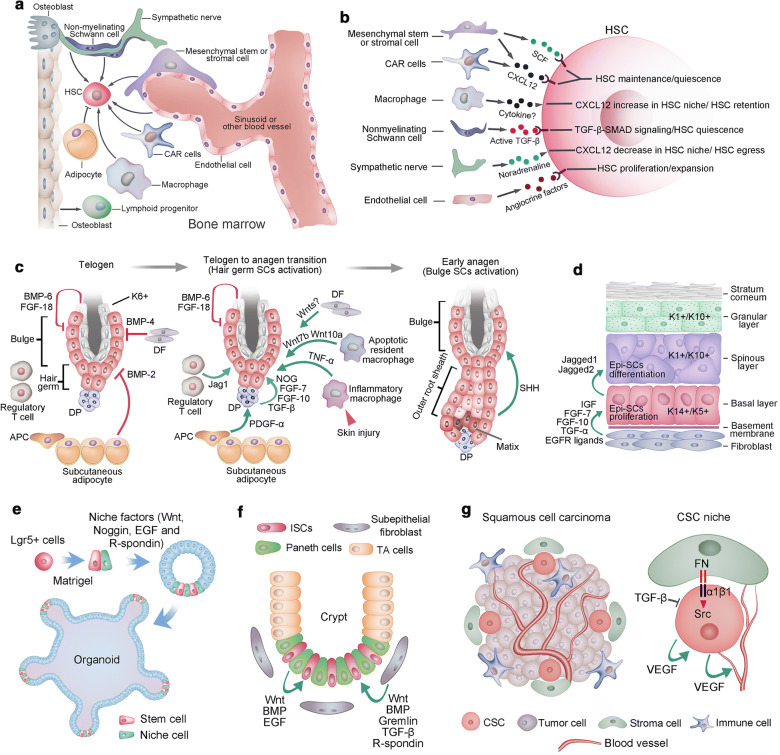
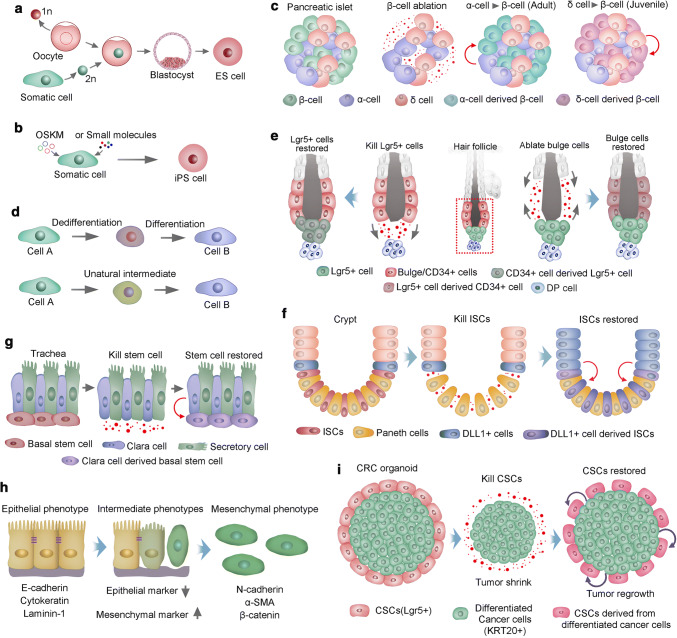
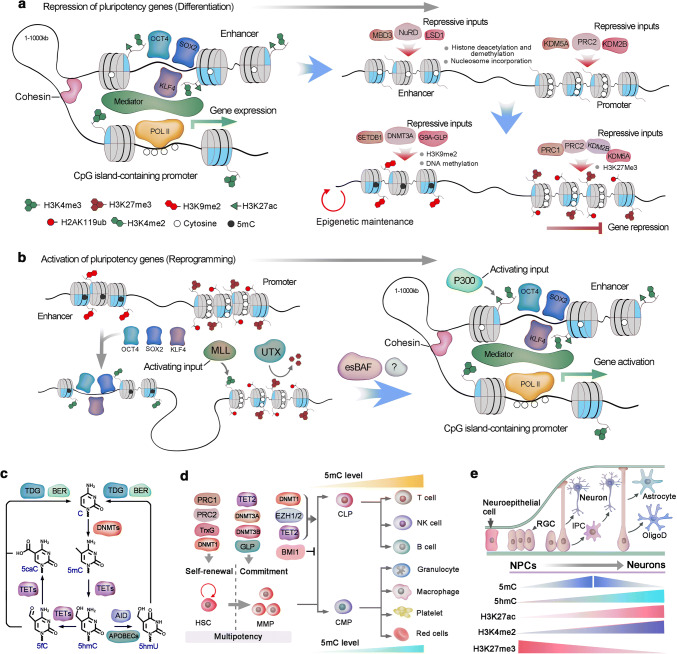
References
-
- Becker AJ, Mc CE, Till JE. Cytological demonstration of the clonal nature of spleen colonies derived from transplanted mouse marrow cells. Nature. 1963;197:452–454. - PubMed
-
- Kretzschmar K, Watt FM. Lineage tracing. Cell. 2012;148(1–2):33–45. - PubMed
-
- Adams GB, Scadden DT. The hematopoietic stem cell in its place. Nat Immunol. 2006;7(4):333–337. - PubMed
-
- de Rooij DG. Proliferation and differentiation of spermatogonial stem cells. Reproduction. 2001;121(3):347–354. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
