

Functionally significant metabolic differences between B and T lymphocyte lineages
- PMID: 31318442
- PMCID: PMC6742768
- DOI: 10.1111/imm.13098
Functionally significant metabolic differences between B and T lymphocyte lineages
Abstract
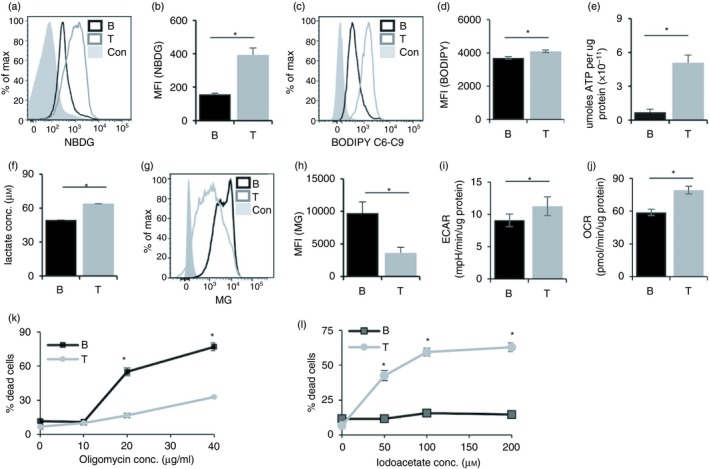
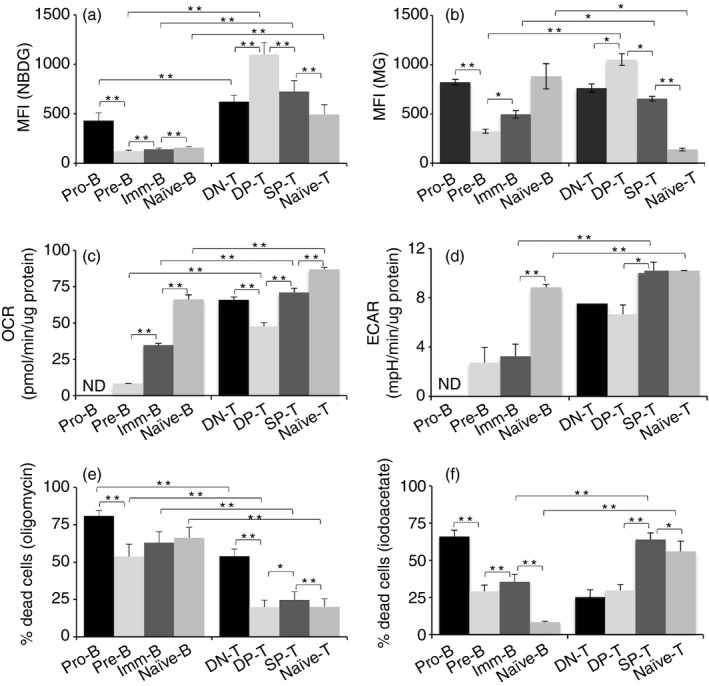
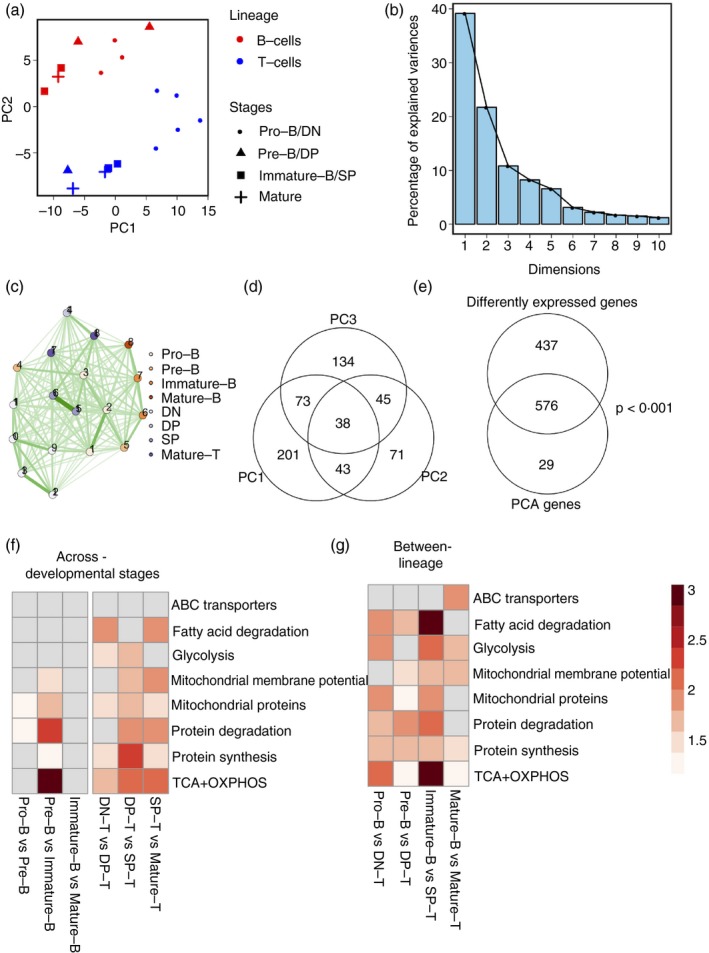
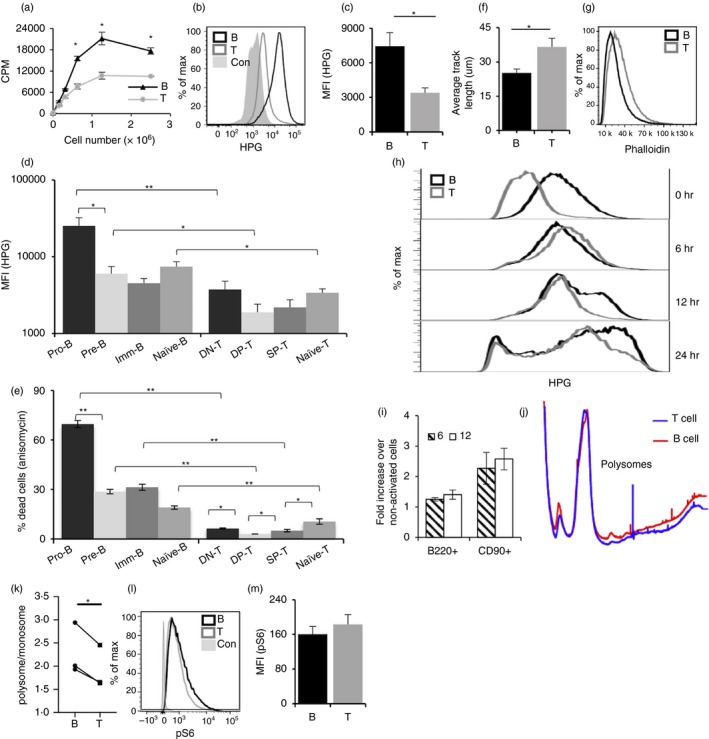
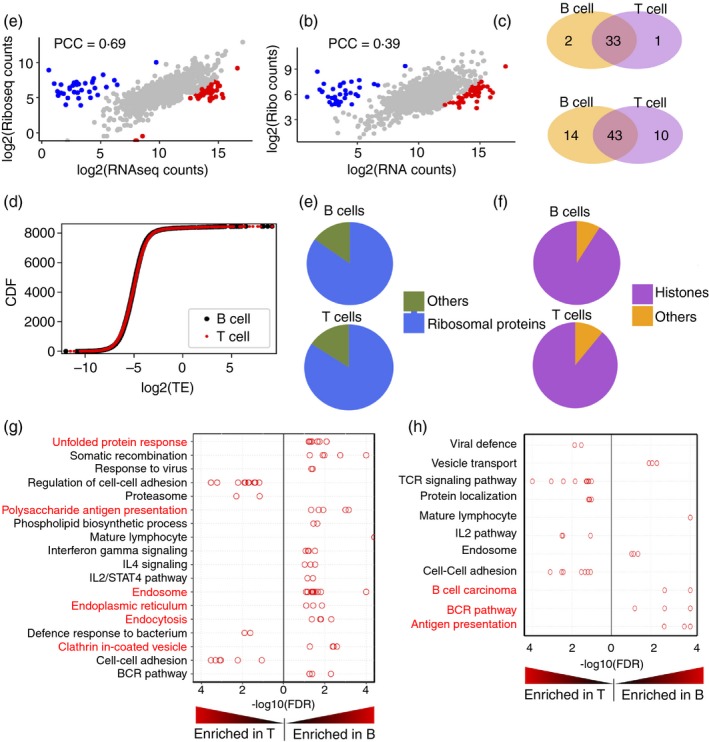
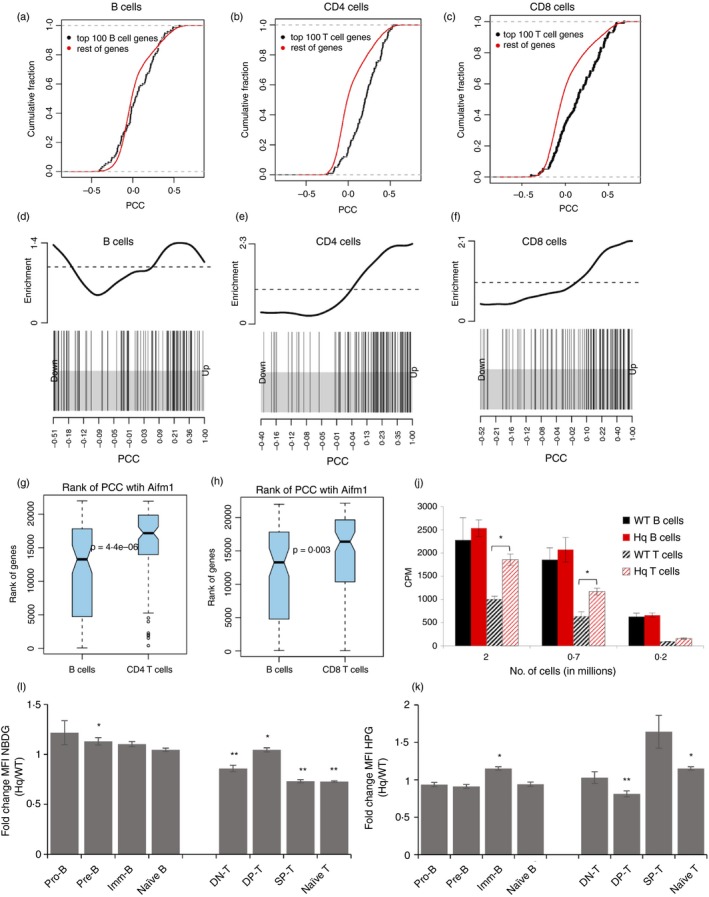
Activation of B and T lymphocytes leads to major remodelling of the metabolic landscape of the cells enabling their post-activation functions. However, naive B and T lymphocytes also show metabolic differences, and the genesis, nature and functional significance of these differences are not yet well understood. Here we show that resting B-cells appeared to have lower energy demands than resting T-cells as they consumed lower levels of glucose and fatty acids and produced less ATP. Resting B-cells are more dependent on OXPHOS, while T-cells show more dependence on aerobic glycolysis. However, despite an apparently higher energy demand, T lineage cells showed lower rates of protein synthesis than equivalent B lineage stages. These metabolic differences between the two lineages were established early during lineage differentiation, and were functionally significant. Higher levels of protein synthesis in B-cells were associated with increased synthesis of MHC class II molecules and other proteins associated with antigen internalization, transport and presentation. The combination of higher energy demand and lower protein synthesis in T-cells was consistent with their higher ATP-dependent motility. Our data provide an integrated perspective of the metabolic differences and their functional implications between the B and T lymphocyte lineages.
Keywords: B-cells; RNA-seq; T-cells; metabolic differences; ribo-seq.
© 2019 John Wiley & Sons Ltd.
Conflict of interest statement
SR is a non‐executive director of Ahammune Biosciences Private Limited, Pune, India, and a member of the scientific advisory boards of Curadev Pharma Private Limited, NOIDA, India, and Mynvax Private Limited, Bangalore, India. Other authors have no financial interests to declare.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
