

Conservation, acquisition, and functional impact of sex-biased gene expression in mammals
- PMID: 31320509
- PMCID: PMC6896219
- DOI: 10.1126/science.aaw7317
Conservation, acquisition, and functional impact of sex-biased gene expression in mammals
Abstract
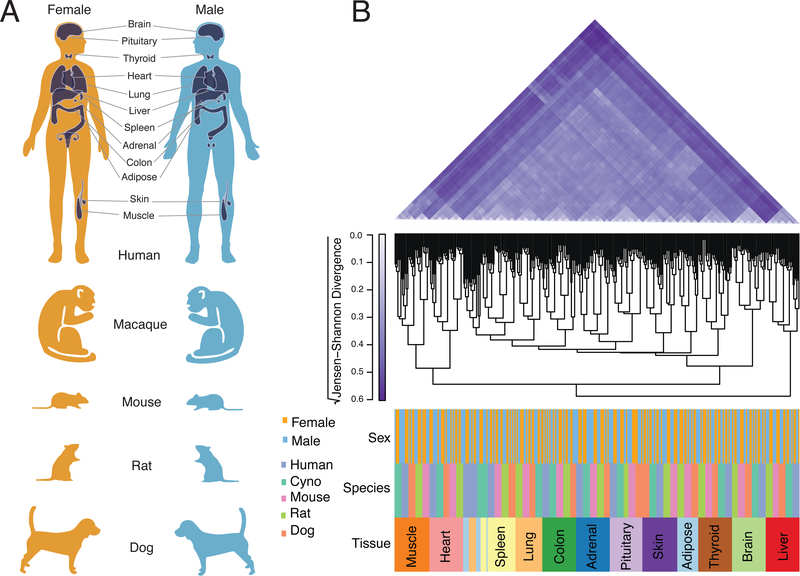
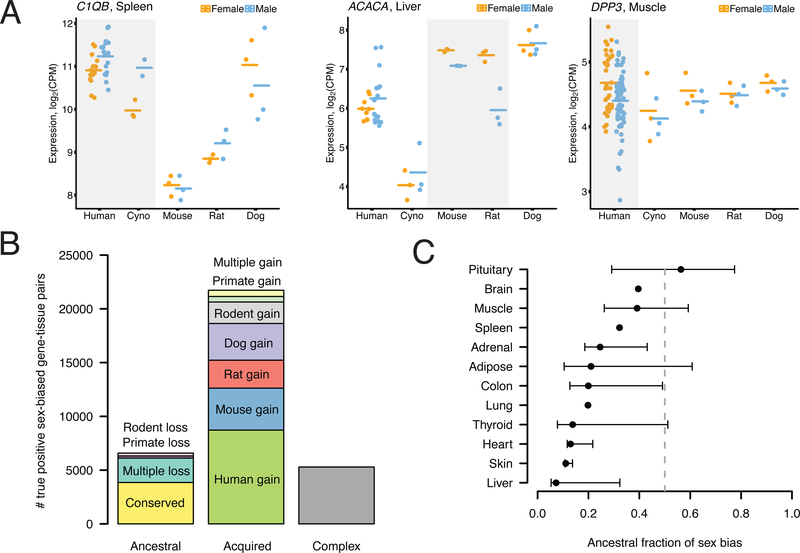
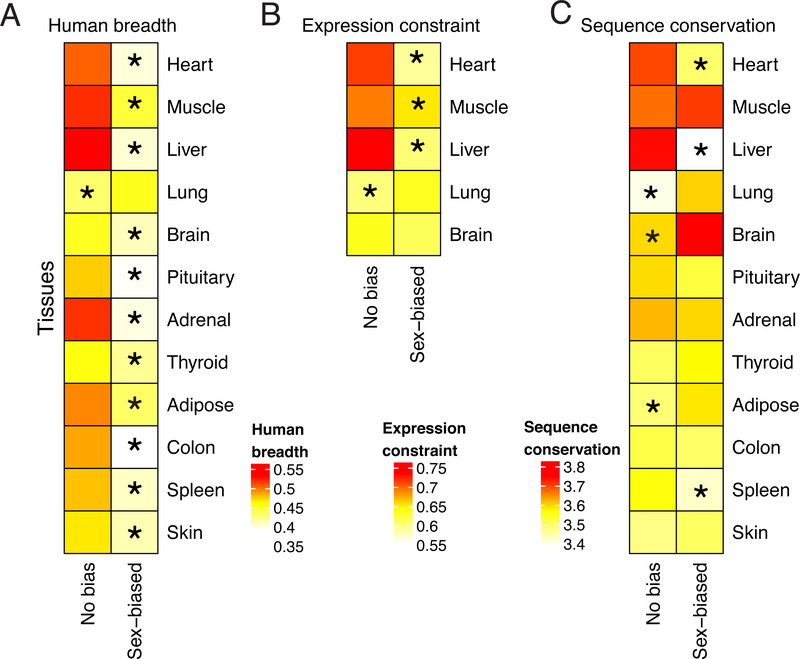
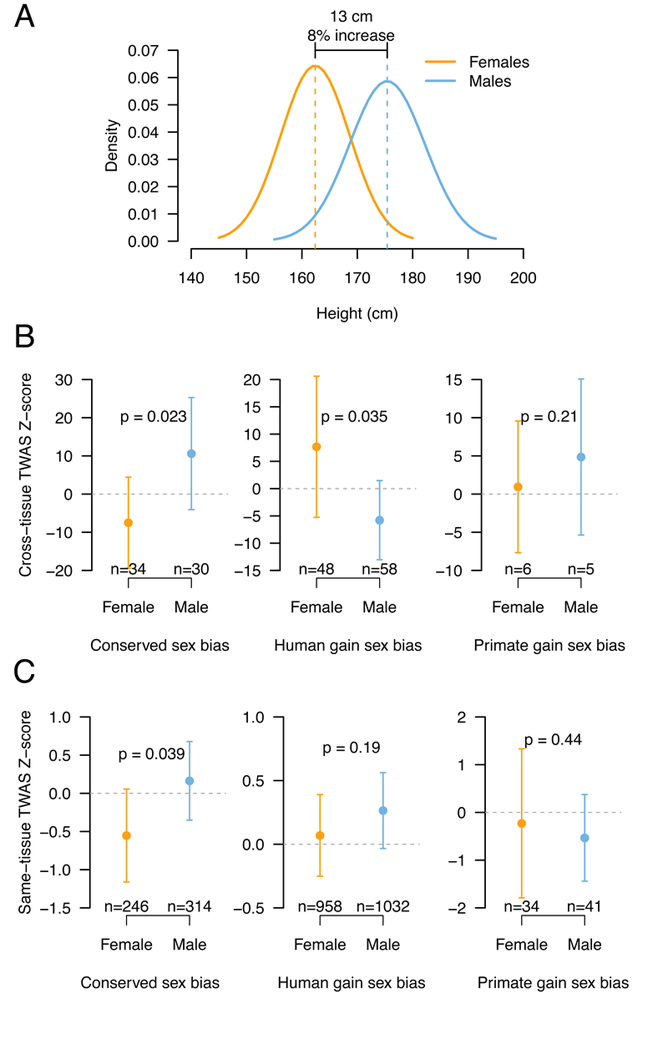
Sex differences abound in human health and disease, as they do in other mammals used as models. The extent to which sex differences are conserved at the molecular level across species and tissues is unknown. We surveyed sex differences in gene expression in human, macaque, mouse, rat, and dog, across 12 tissues. In each tissue, we identified hundreds of genes with conserved sex-biased expression-findings that, combined with genomic analyses of human height, explain ~12% of the difference in height between females and males. We surmise that conserved sex biases in expression of genes otherwise operating equivalently in females and males contribute to sex differences in traits. However, most sex-biased expression arose during the mammalian radiation, which suggests that careful attention to interspecies divergence is needed when modeling human sex differences.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures

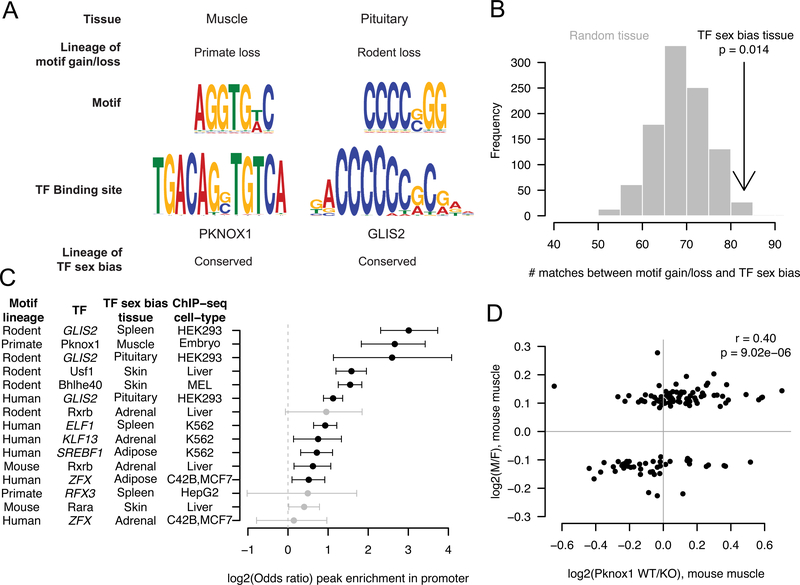
References
-
- Wells JCK, Sexual dimorphism of body composition. Best Pract. Res. Clin. Endocrinol. Metab 21, 415–430 (2007). - PubMed
-
- Green HJ, Fraser IG, Ranney DA, Male and female differences in enzyme activities of energy metabolism in vastus lateralis muscle. J. Neurol. Sci 65, 323–331 (1984). - PubMed
-
- Klein SL, Flanagan KL, Sex differences in immune responses. Nat. Rev. Immunol 16, 626–638 (2016). - PubMed
-
- Hayward CS, Kalnins WV, Kelly RP, Gender-related differences in left ventricular chamber function. Cardiovasc. Res 49, 340–350 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
