

Co-option of neurotransmitter signaling for inter-organismal communication in C. elegans
- PMID: 31320626
- PMCID: PMC6639374
- DOI: 10.1038/s41467-019-11240-7
Co-option of neurotransmitter signaling for inter-organismal communication in C. elegans
Abstract
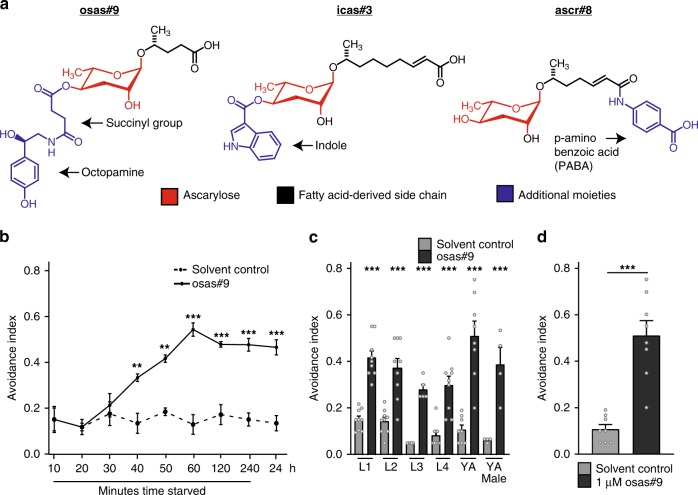
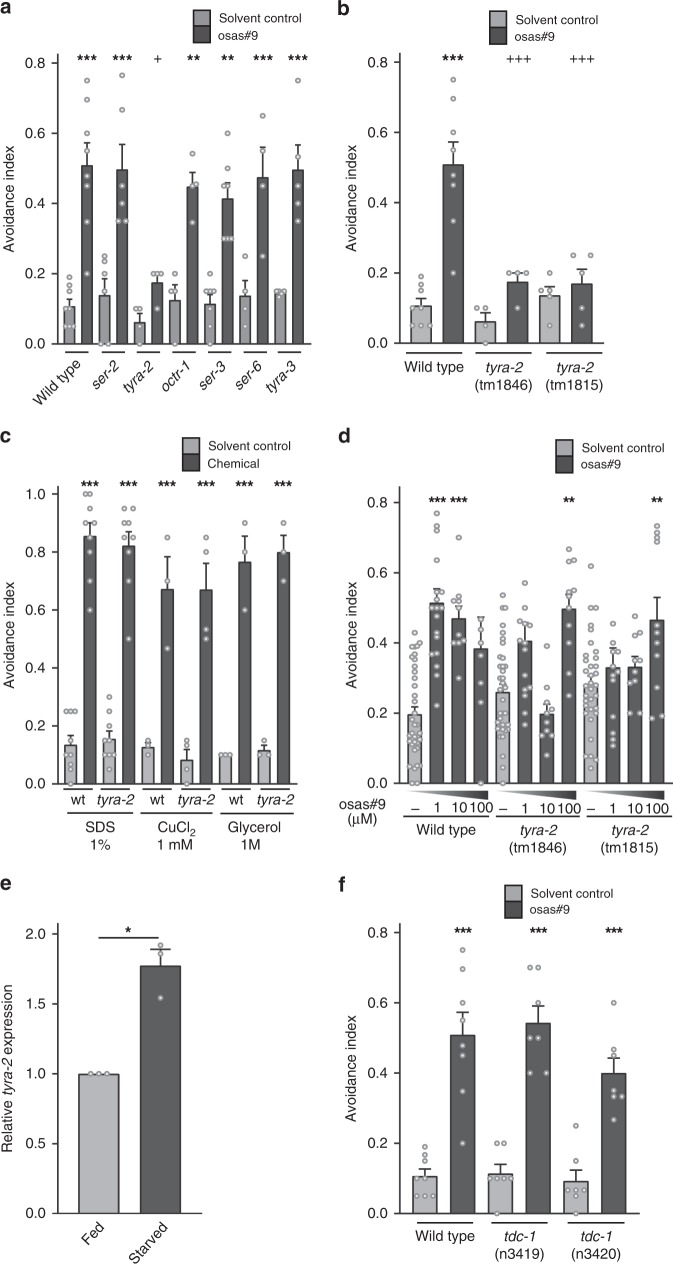
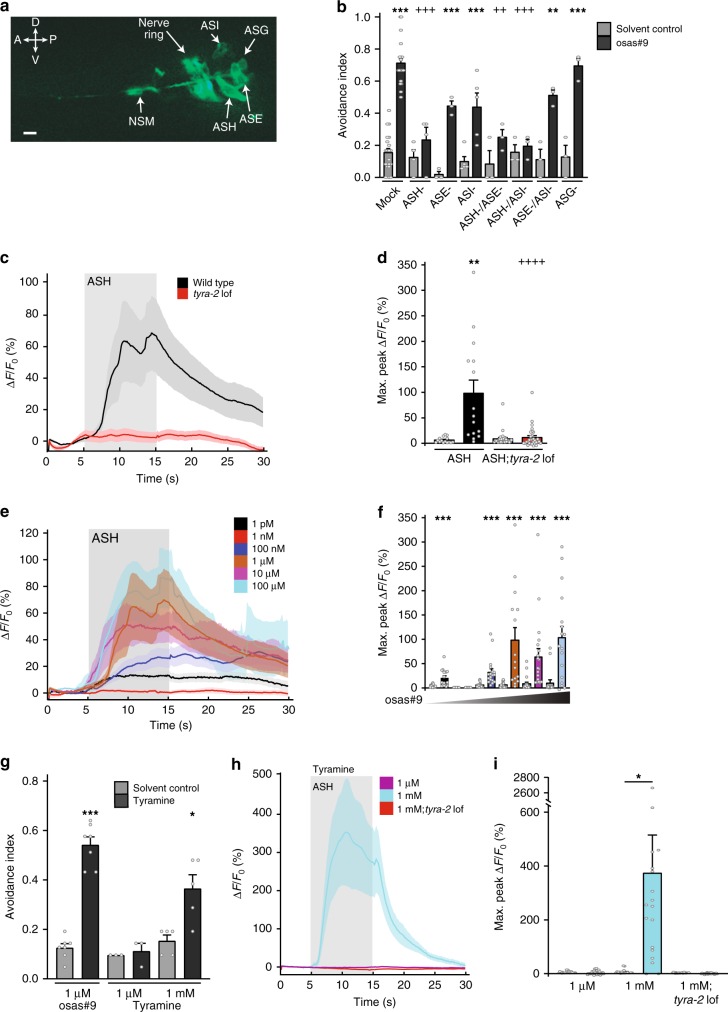
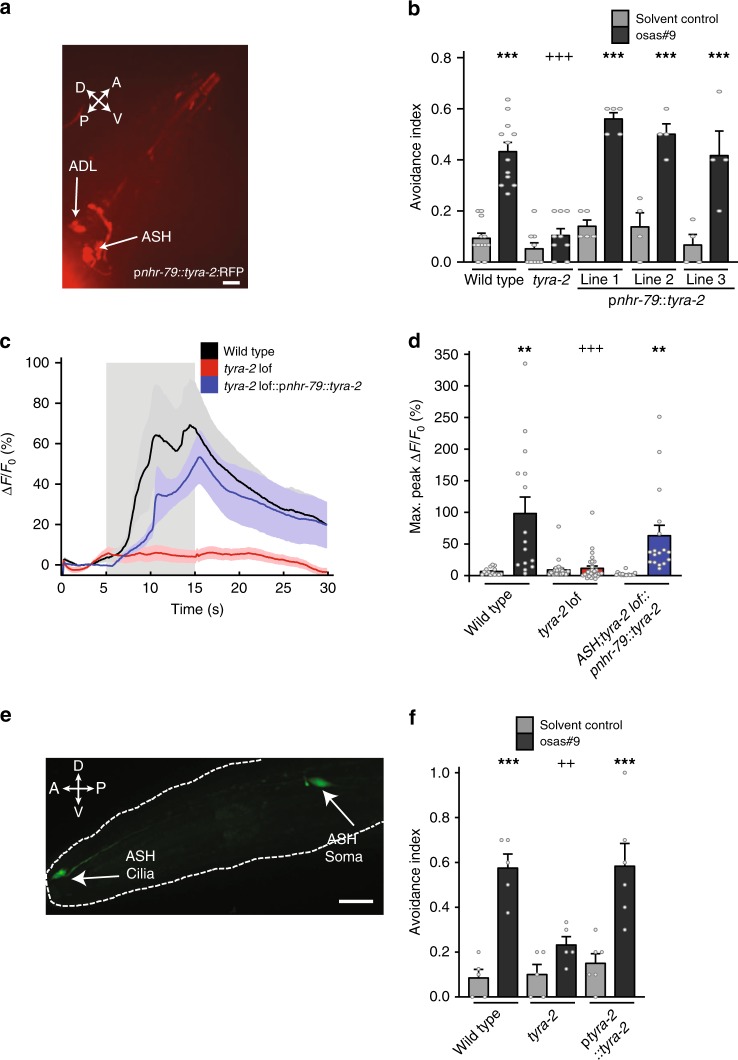
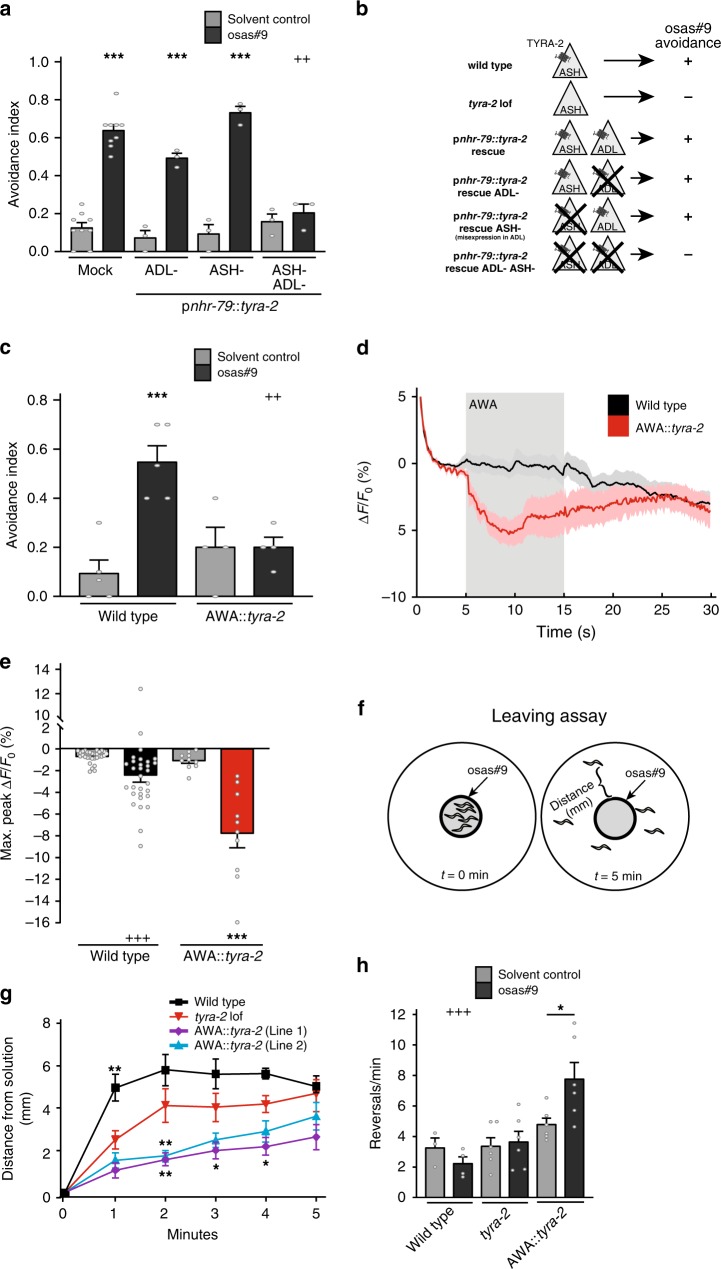
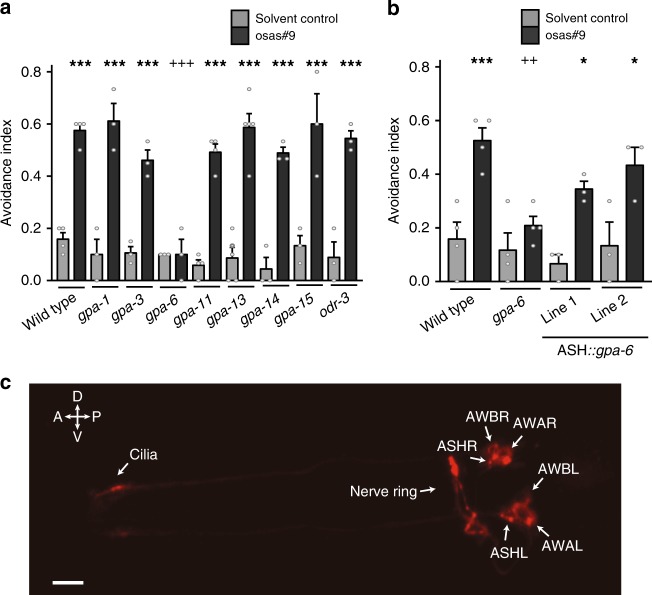
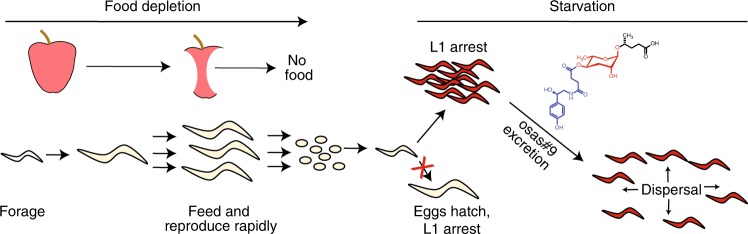
Biogenic amine neurotransmitters play a central role in metazoan biology, and both their chemical structures and cognate receptors are evolutionarily conserved. Their primary roles are in cell-to-cell signaling, as biogenic amines are not normally recruited for communication between separate individuals. Here, we show that in the nematode C. elegans, a neurotransmitter-sensing G protein-coupled receptor, TYRA-2, is required for avoidance responses to osas#9, an ascaroside pheromone that incorporates the neurotransmitter, octopamine. Neuronal ablation, cell-specific genetic rescue, and calcium imaging show that tyra-2 expression in the nociceptive neuron, ASH, is necessary and sufficient to induce osas#9 avoidance. Ectopic expression in the AWA neuron, which is generally associated with attractive responses, reverses the response to osas#9, resulting in attraction instead of avoidance behavior, confirming that TYRA-2 partakes in the sensing of osas#9. The TYRA-2/osas#9 signaling system represents an inter-organismal communication channel that evolved via co-option of a neurotransmitter and its cognate receptor.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Wilson, E. O. Sociobiology (The Belknap, 1980).
-
- Wyatt, T. D. Pheromones And Animal Behavior: Chemical Signals And Signatures. (Cambridge Univ. Press, 2014).
-
- Roschina, V. V. Microbial Endocrnology p. 17–52 (Springer New York, 2010).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
