

Alterations in Organismal Physiology, Impaired Stress Resistance, and Accelerated Aging in Drosophila Flies Adapted to Multigenerational Proteome Instability
- PMID: 31320986
- PMCID: PMC6610734
- DOI: 10.1155/2019/7823285
Alterations in Organismal Physiology, Impaired Stress Resistance, and Accelerated Aging in Drosophila Flies Adapted to Multigenerational Proteome Instability
Abstract
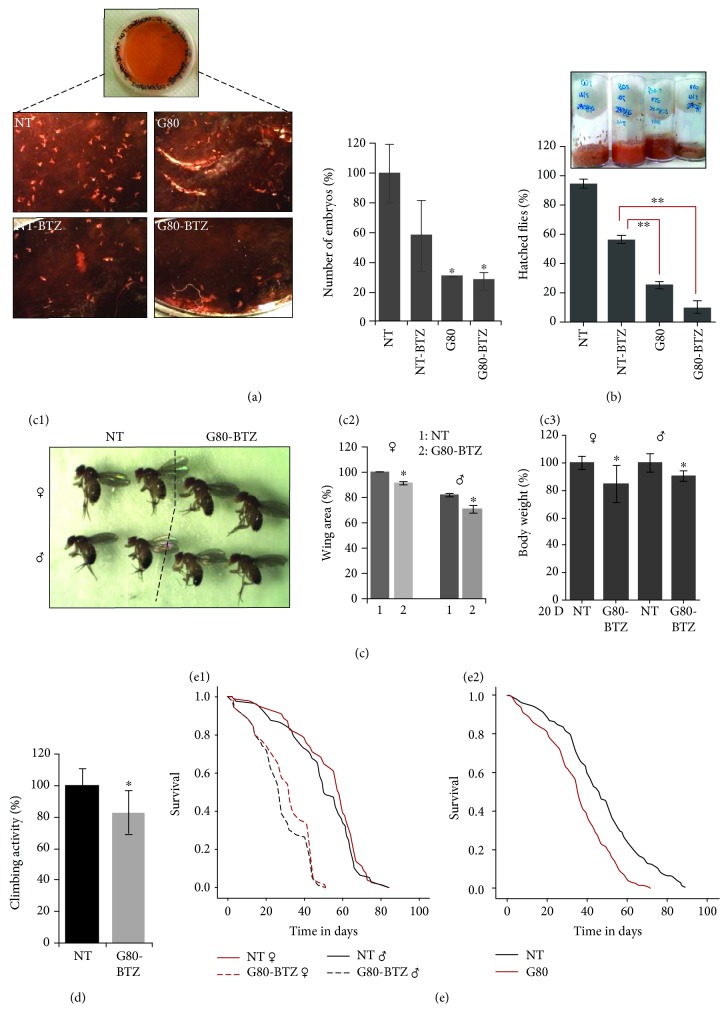
Being an assembly of highly sophisticated protein machines, cells depend heavily on proteostatic modules functionality and on adequate supply of energetic molecules for maintaining proteome stability. Yet, our understanding of the adaptations induced by multigenerational proteotoxic stress is limited. We report here that multigenerational (>80 generations) proteotoxic stress in OregonR flies induced by constant exposure to developmentally nonlethal doses of the proteasome inhibitor bortezomib (BTZ) (G80-BTZ flies) increased proteome instability and redox imbalance, reduced fecundity and body size, and caused neuromuscular defects; it also accelerated aging. G80-BTZ flies were mildly resistant to increased doses of BTZ and showed no age-related loss of proteasome activity; these adaptations correlated with sustained upregulation of proteostatic modules, which however occurred at the cost of minimal responses to increased BTZ doses and increased susceptibility to various types of additional proteotoxic stress, namely, autophagy inhibition or thermal stress. Multigenerational proteome instability and redox imbalance also caused metabolic reprogramming being evidenced by altered mitochondrial biogenesis and suppressed insulin/IGF-like signaling (IIS) in G80-BTZ flies. The toxic effects of multigenerational proteome instability could be partially mitigated by a low-protein diet that extended G80-BTZ flies' longevity. Overall, persistent proteotoxic stress triggers a highly conserved adaptive metabolic response mediated by the IIS pathway, which reallocates resources from growth and longevity to somatic preservation and stress tolerance. Yet, these trade-off adaptations occur at the cost of accelerated aging and/or reduced tolerance to additional stress, illustrating the limited buffering capacity of survival pathways.
Figures





References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
