

Nonsense-mediated mRNA decay of the ferric and cupric reductase mRNAs FRE1 and FRE2 in Saccharomyces cerevisiae
- PMID: 31322728
- PMCID: PMC6878129
- DOI: 10.1002/1873-3468.13545
Nonsense-mediated mRNA decay of the ferric and cupric reductase mRNAs FRE1 and FRE2 in Saccharomyces cerevisiae
Abstract
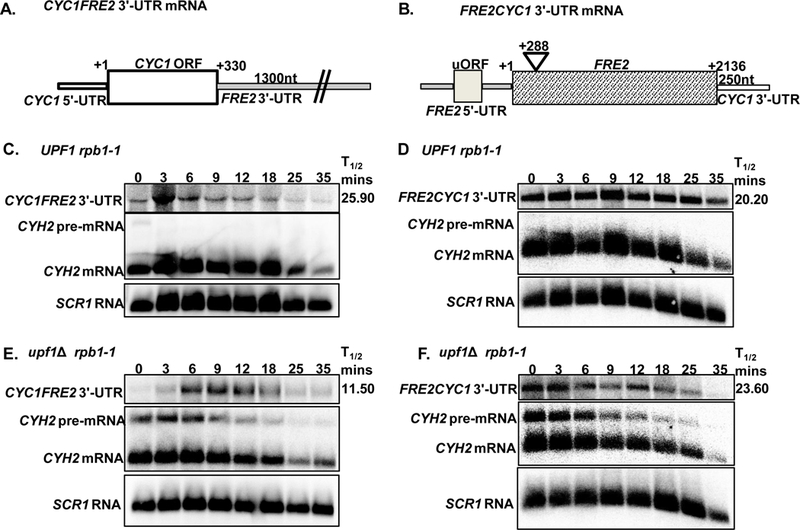
The nonsense-mediated mRNA decay (NMD) pathway regulates mRNAs that aberrantly terminate translation. This includes aberrant mRNAs and functional natural mRNAs. Natural mRNA degradation by NMD is triggered by mRNA features and environmental cues. Saccharomyces cerevisiae encodes multiple proteins with ferric and cupric reductase activity. Here, we examined the regulation by NMD of two mRNAs, FRE1 and FRE2, encoding ferric and cupric reductases in S. cerevisiae. We found that FRE2 mRNAs are regulated by NMD under noninducing conditions and that the FRE2 3'-UTR contributes to the degradation of the mRNAs by NMD. Conversely, FRE1 mRNAs are not regulated by NMD under comparable conditions. These findings suggest that regulation of functionally related mRNAs by NMD can be differential and conditional.
Keywords: FRE1-2 mRNA; Saccharomyces cerevisiae; biometal homeostasis; mRNA decay; nonsense-mediated mRNA decay.
© 2019 Federation of European Biochemical Societies.
Figures



References
-
- He F, Li X, Spatrick P, Casillo R, Dong S, and Jacobson A (2003). Genome-wide analysis of mRNAs regulated by the nonsense-mediated and 5’ to 3’ mRNA decay pathways in yeast. Mol Cell 12, 1439–1452. - PubMed
-
- Mendell JT, Sharifi NA, Meyers JL, Martinez-Murillo F, and Dietz HC (2004). Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and mutes genomic noise. Nature genetics 36, 1073–1078. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
