

Chronic adolescent exposure to ∆9-tetrahydrocannabinol decreases NMDA current and extrasynaptic plasmalemmal density of NMDA GluN1 subunits in the prelimbic cortex of adult male mice
- PMID: 31323660
- PMCID: PMC6901492
- DOI: 10.1038/s41386-019-0466-9
Chronic adolescent exposure to ∆9-tetrahydrocannabinol decreases NMDA current and extrasynaptic plasmalemmal density of NMDA GluN1 subunits in the prelimbic cortex of adult male mice
Abstract
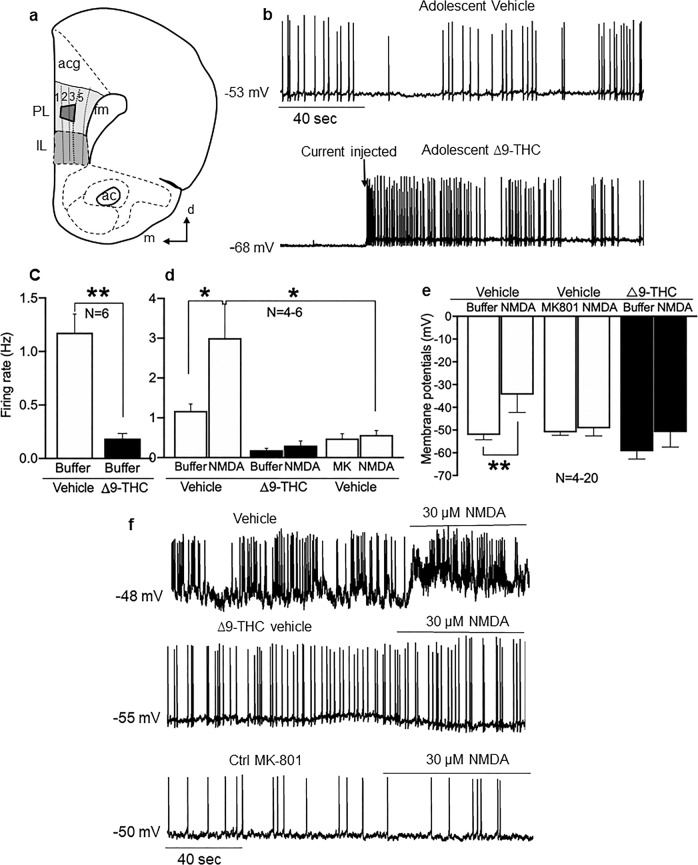
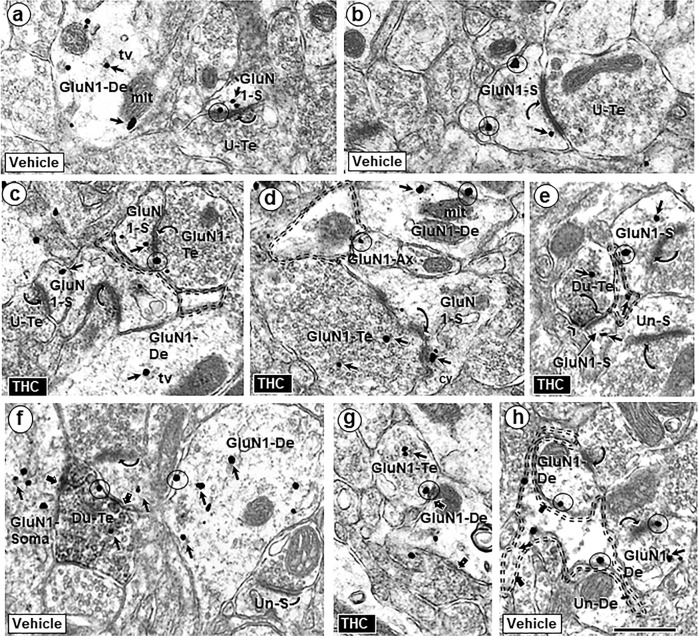
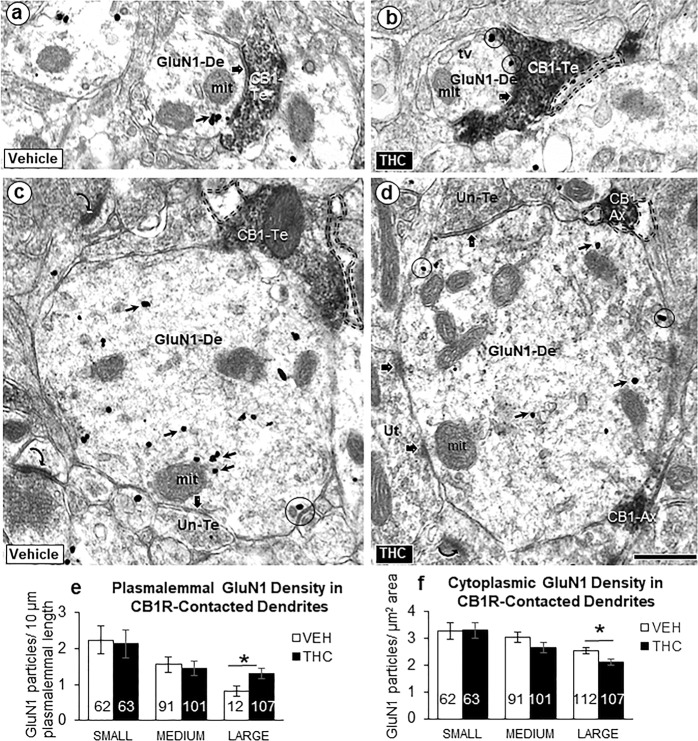
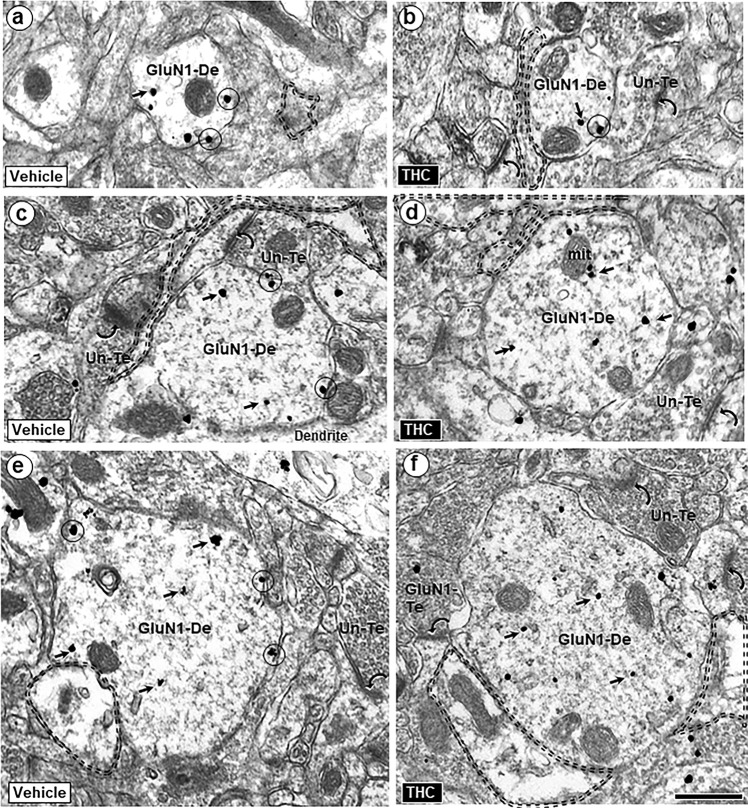
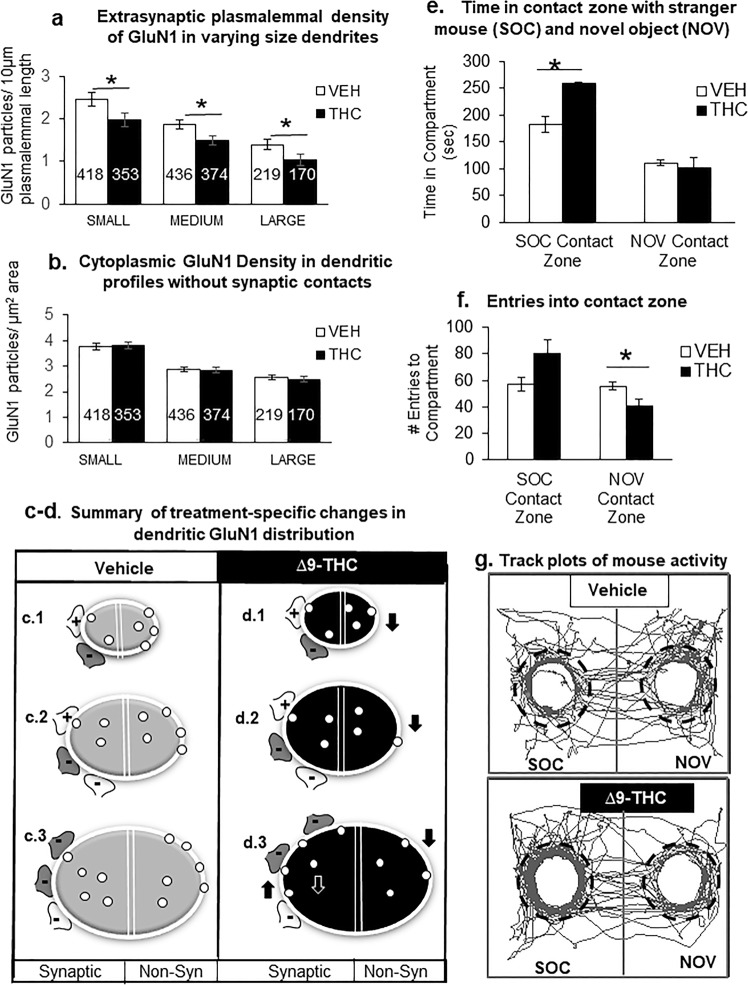
Adolescence is a vulnerable period of development when limbic connection of the prefrontal cortex (PFC) involved in emotional processing may be rendered dysfunctional by chronic exposure to delta-9-tetrahydrocannabinol (∆9-THC), the major psychoactive compound in marijuana. Cannabinoid-1 receptors (CB1Rs) largely mediate the central neural effects of ∆9-THC and endocannabinoids that regulate NMDA receptor-dependent synaptic plasticity of glutamatergic synapses in the prelimbic prefrontal cortex (PL-PFC). Thus, chronic occupancy of CB1Rs by ∆9-THC during adolescence may competitively decrease the functional expression and activity of NMDA receptors in the mature PL-PFC. We used a multidisciplinary approach to test this hypothesis in adult C57BL/6J male mice that received vehicle or ∆9-THC in escalating doses (2.5-10 mg/kg/ip) through adolescence (postnatal day 29-43). In comparison with vehicle, the mice receiving ∆9-THC showed a hyperpolarized resting membrane potential, decreased spontaneous firing rate, increased current-induced firing threshold, and decreased depolarizing response to NMDA in deep-layer PL-PFC neurons analyzed by current-clamp recordings. Electron microscopic immunolabeling in the PL-PFC of adult mice that had received Δ9-THC only during adolescence showed a significant (1) decrease in the extrasynaptic plasmalemmal density of obligatory GluN1-NMDA subunits in dendrites of all sizes and (2) a shift from cytoplasmic to plasmalemmal distribution of GluN1 in large dendrites receiving mainly inhibitory-type synapses from CB1R-labeled terminals. From these results and concomitant behavioral studies, we conclude that social dysfunctions resulting from excessive intake of ∆9-THC in the increasingly available marijuana products used by male teens may largely reflect circuit defects in PL-PFC networks communicating through endocannabinoid-regulated NMDA receptors.
Figures
References
-
- Hashimotodani Y, Ohno-Shosaku T, Kano M. Endocannabinoids and synaptic function in the CNS. Neuroscientist. 2007;13:127–37. - PubMed
-
- Fortin DA, Levine ES. Differential effects of endocannabinoids on glutamatergic and GABAergic inputs to layer 5 pyramidal neurons. Cereb Cortex. 2007;17:163–74. - PubMed
-
- Auclair N, Otani S, Soubrie P, Crepel F. Cannabinoids modulate synaptic strength and plasticity at glutamatergic synapses of rat prefrontal cortex pyramidal neurons. J Neurophysiol. 2000;83:3287–93. - PubMed
-
- Murphy M, Mills S, Winstone J, Leishman E, Wager-Miller J, Bradshaw H, et al. Chronic adolescent delta(9)-tetrahydrocannabinol treatment of male mice leads to long-term cognitive and behavioral dysfunction, which are prevented by concurrent cannabidiol treatment. Cannabis Cannabinoid Res. 2017;2:235–46. - PMC - PubMed
-
- Dow-Edwards D, Silva L. Endocannabinoids in brain plasticity: cortical maturation, HPA axis function and behavior. Brain Res. 2017;1654(Pt B):157–64. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
