

Dopamine Deficiency Reduces Striatal Cholinergic Interneuron Function in Models of Parkinson's Disease
- PMID: 31324539
- PMCID: PMC7102938
- DOI: 10.1016/j.neuron.2019.06.013
Dopamine Deficiency Reduces Striatal Cholinergic Interneuron Function in Models of Parkinson's Disease
Abstract
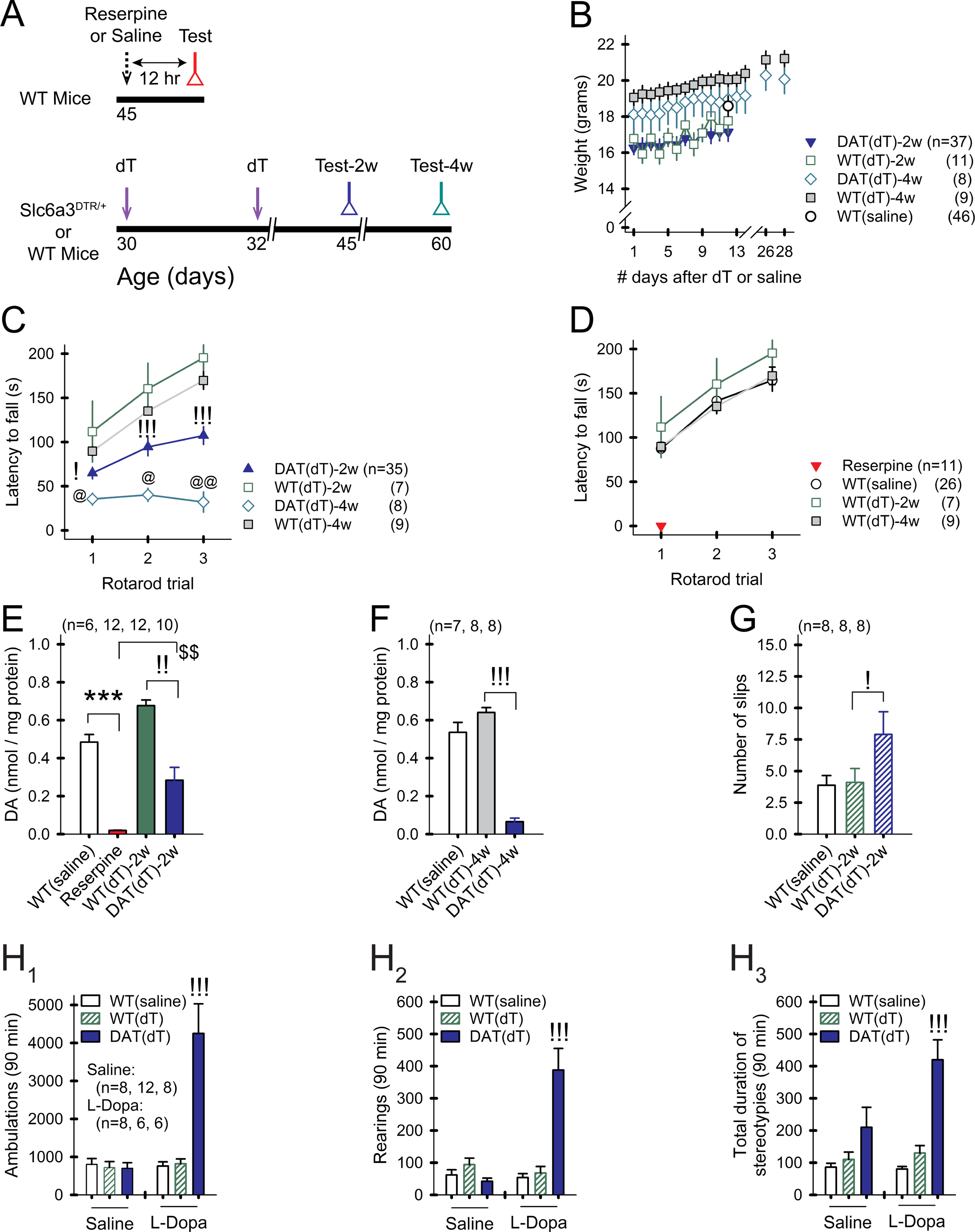
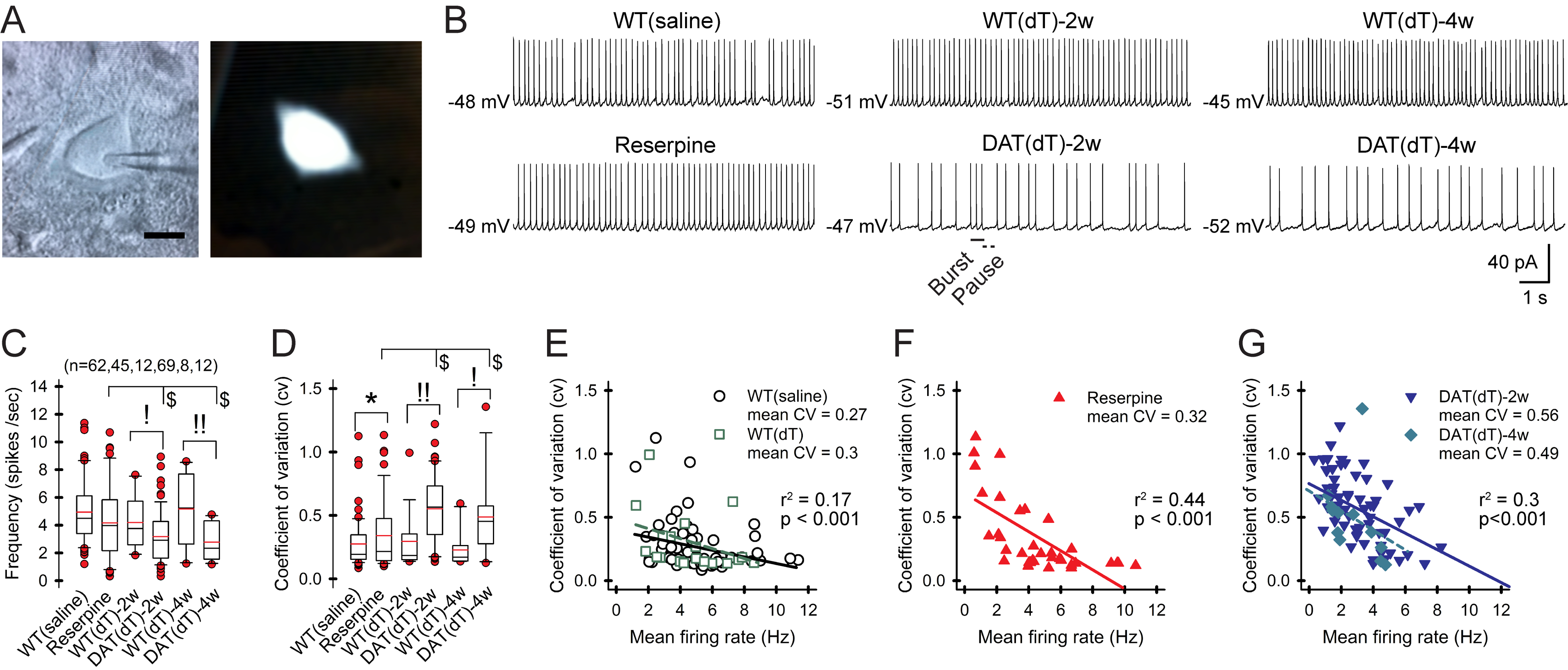
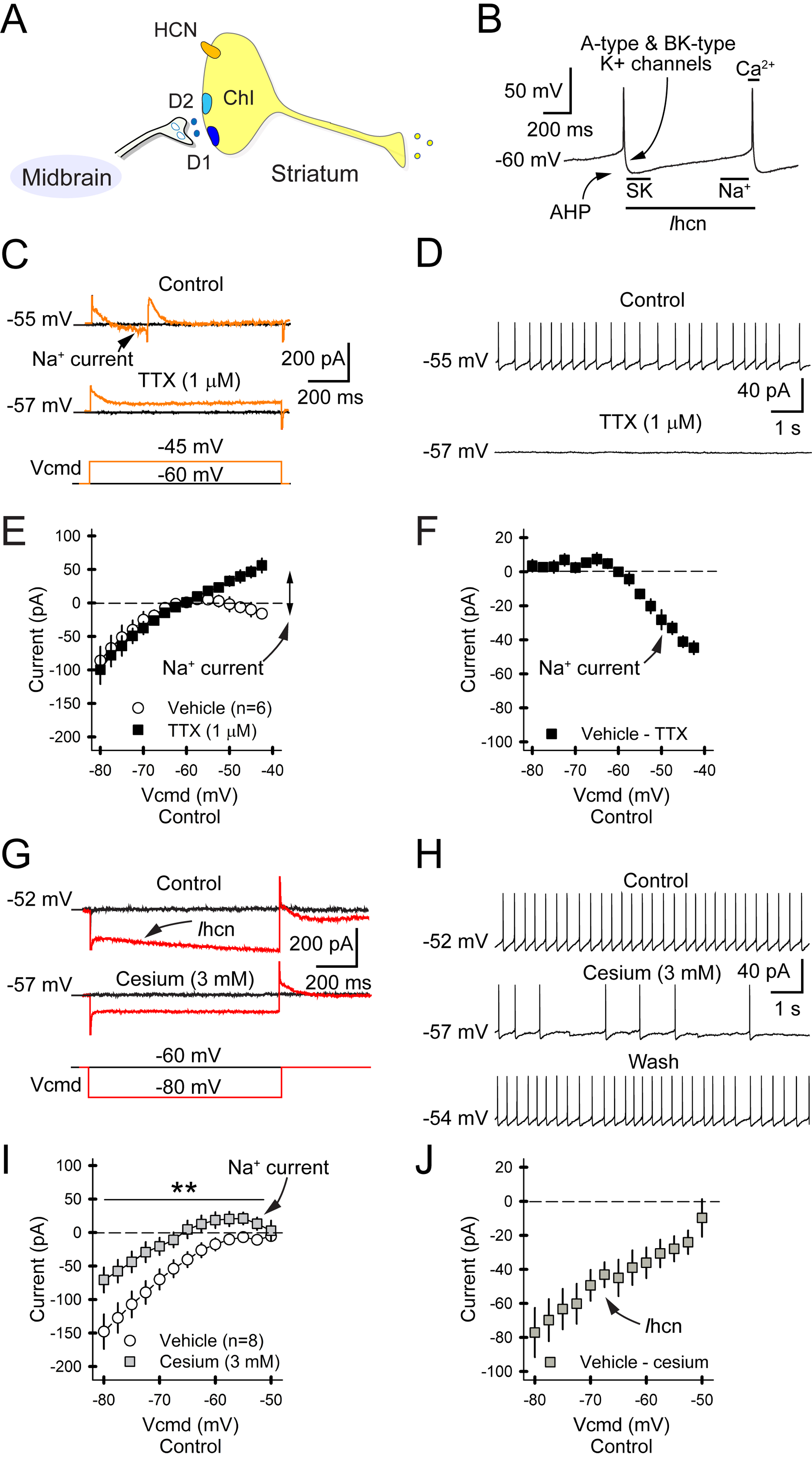
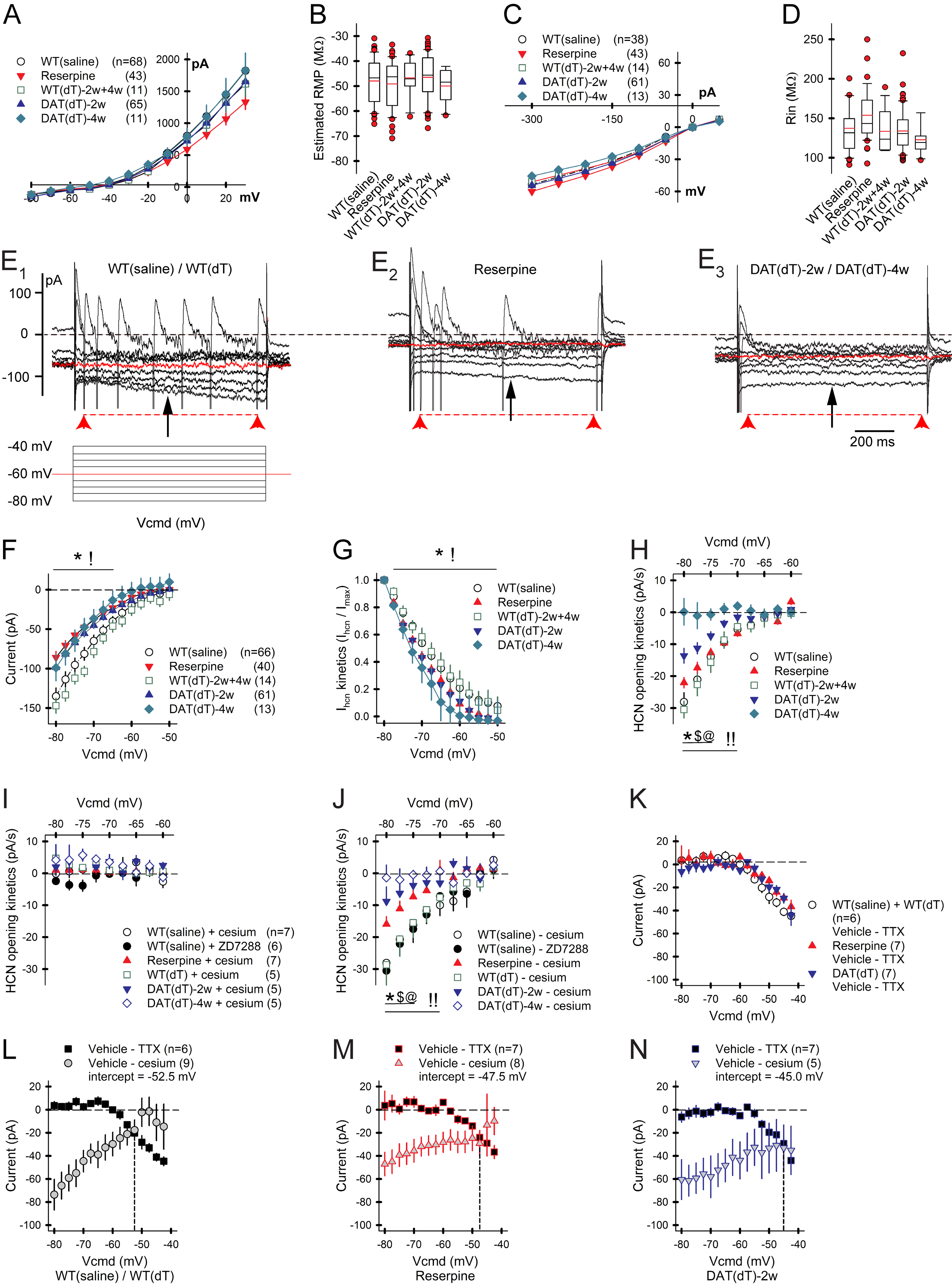
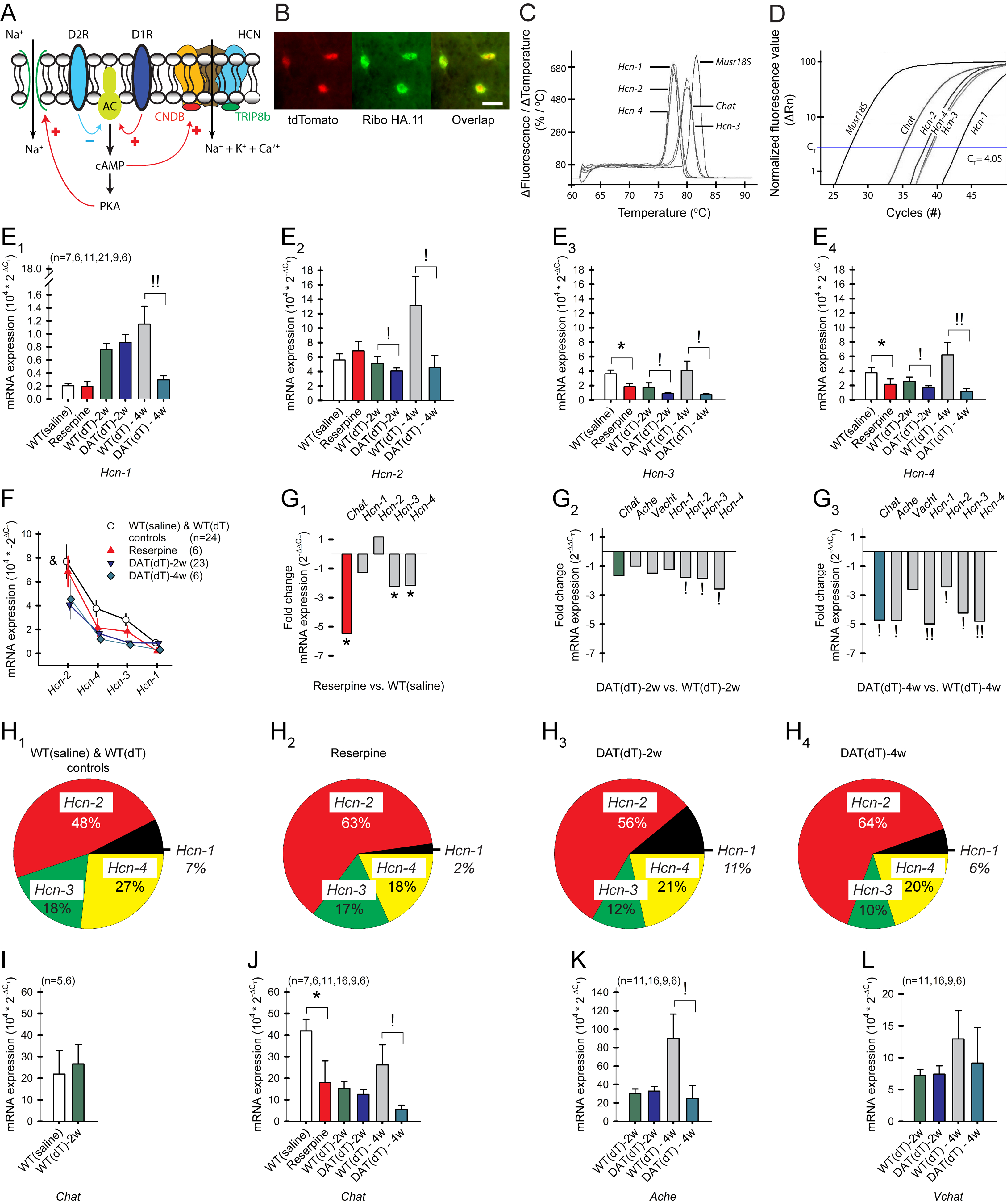
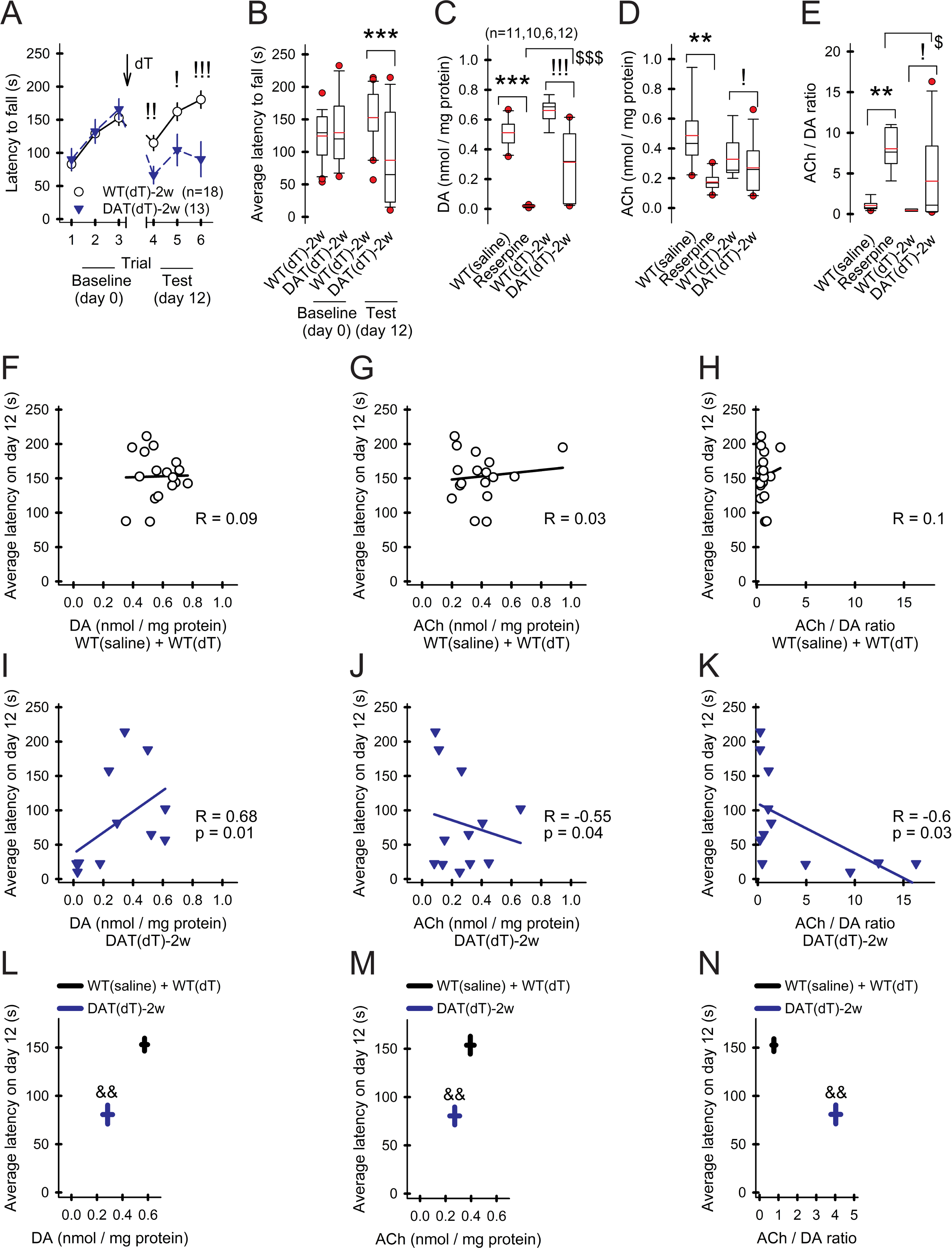
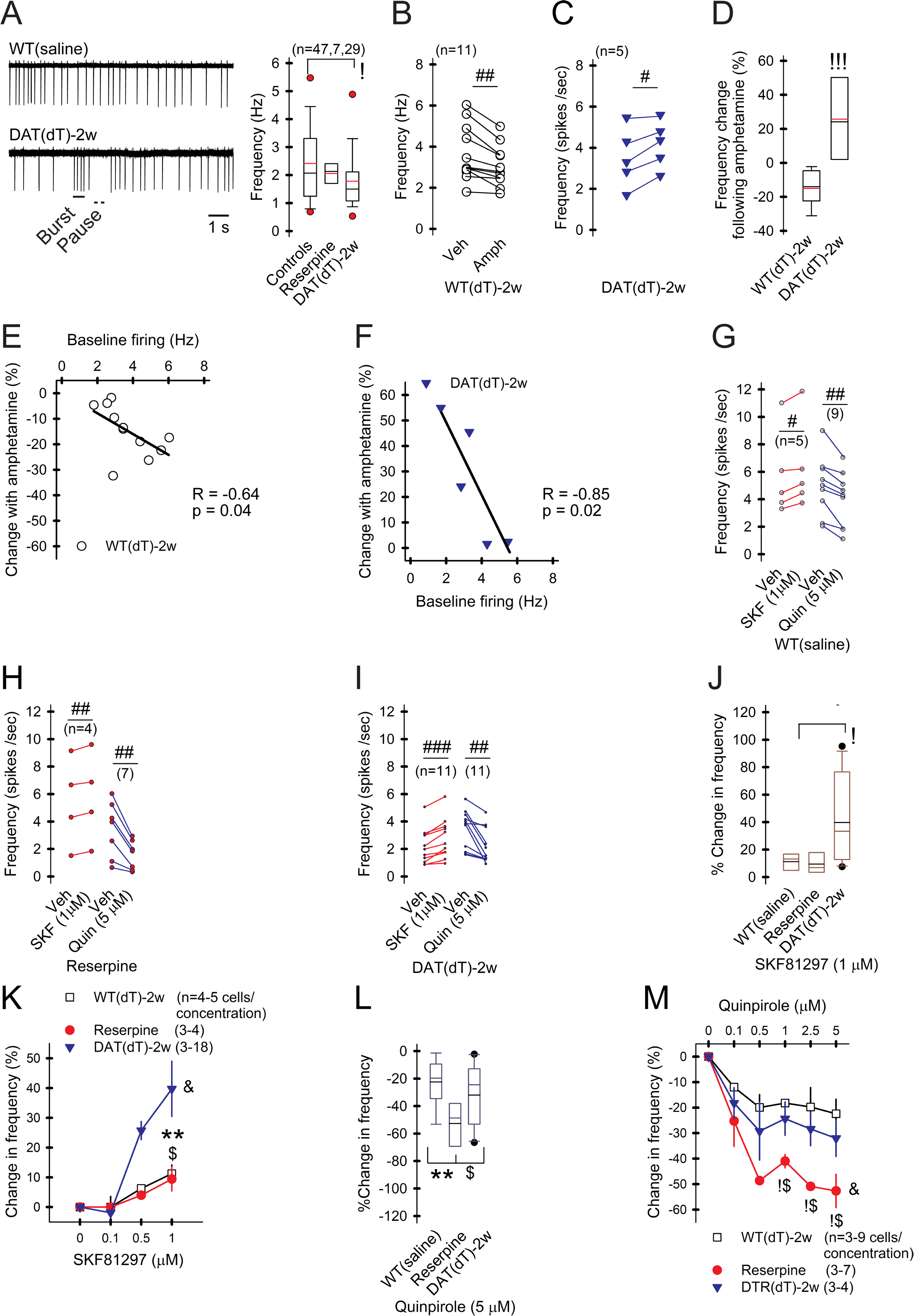
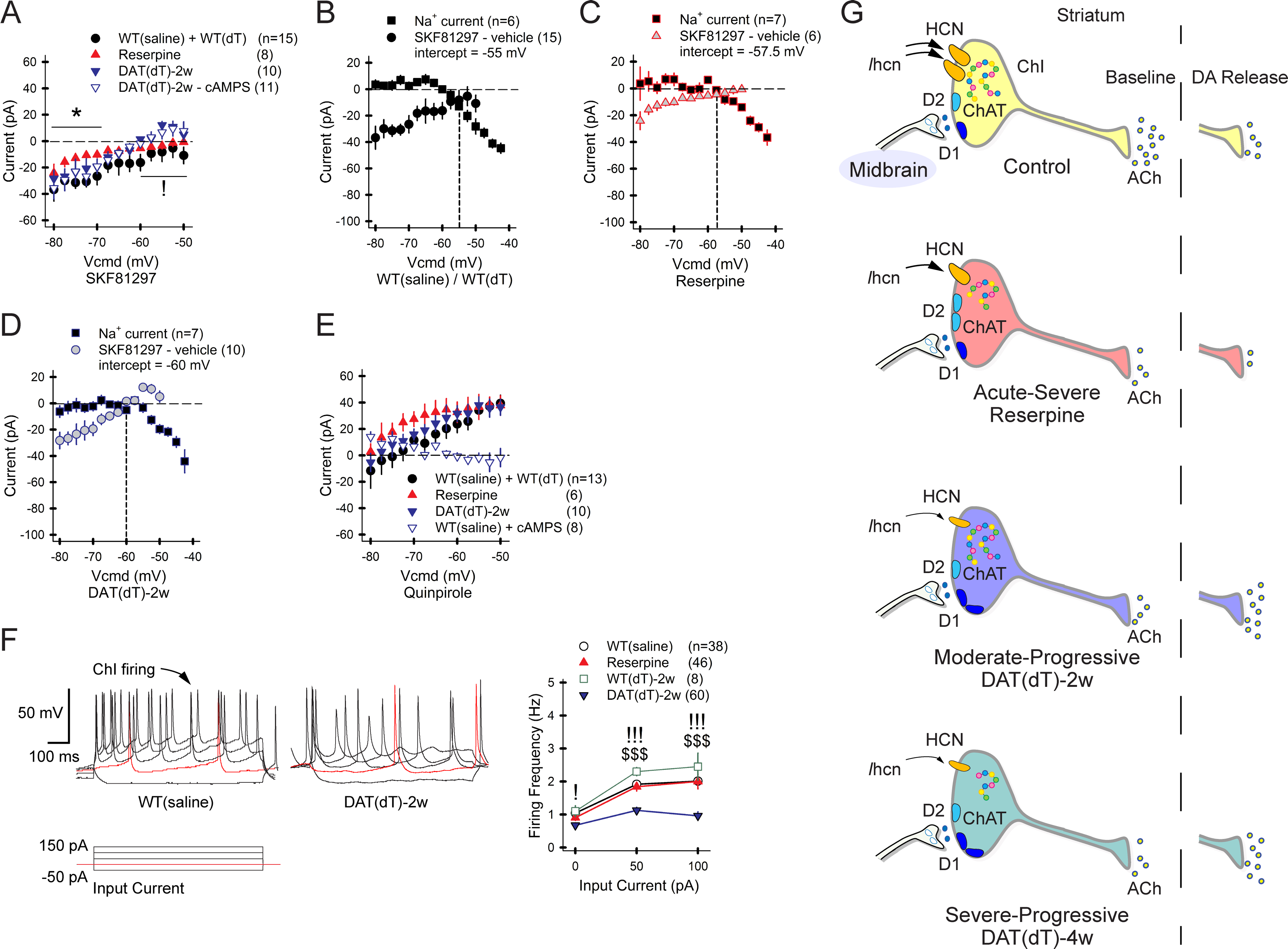
Motor and cognitive functions depend on the coordinated interactions between dopamine (DA) and acetylcholine (ACh) at striatal synapses. Increased ACh availability was assumed to accompany DA deficiency based on the outcome of pharmacological treatments and measurements in animals that were critically depleted of DA. Using Slc6a3DTR/+ diphtheria-toxin-sensitive mice, we demonstrate that a progressive and L-dopa-responsive DA deficiency reduces ACh availability and the transcription of hyperpolarization-activated cation (HCN) channels that encode the spike timing of ACh-releasing tonically active striatal interneurons (ChIs). Although the production and release of ACh and DA are reduced, the preponderance of ACh over DA contributes to the motor deficit. The increase in striatal ACh relative to DA is heightened via D1-type DA receptors that activate ChIs in response to DA release from residual axons. These results suggest that stabilizing the expression of HCN channels may improve ACh-DA reciprocity and motor function in Parkinson's disease (PD). VIDEO ABSTRACT.
Keywords: HCN channel; RiboTag; acetylcholine; diphtheria toxin; dopamine acetylcholine ratio; dopamine depletion; electrophysiology; receptor.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECELRATION OF INTERESTS
The authors declare no competing interests.
Figures
Similar articles
-
Strength of cholinergic tone dictates the polarity of dopamine D2 receptor modulation of striatal cholinergic interneuron excitability in DYT1 dystonia.Exp Neurol. 2017 Sep;295:162-175. doi: 10.1016/j.expneurol.2017.06.005. Epub 2017 Jun 3. Exp Neurol. 2017. PMID: 28587876 Free PMC article.
-
Striatal muscarinic receptors promote activity dependence of dopamine transmission via distinct receptor subtypes on cholinergic interneurons in ventral versus dorsal striatum.J Neurosci. 2010 Mar 3;30(9):3398-408. doi: 10.1523/JNEUROSCI.5620-09.2010. J Neurosci. 2010. PMID: 20203199 Free PMC article.
-
Contribution of cholinergic interneurons to striatal pathophysiology in Parkinson's disease.Neurochem Int. 2019 Jun;126:1-10. doi: 10.1016/j.neuint.2019.02.019. Epub 2019 Feb 27. Neurochem Int. 2019. PMID: 30825602 Review.
-
Dopaminergic Control of Striatal Cholinergic Interneurons Underlies Cocaine-Induced Psychostimulation.Cell Rep. 2020 Apr 21;31(3):107527. doi: 10.1016/j.celrep.2020.107527. Cell Rep. 2020. PMID: 32320647
-
A new outlook on cholinergic interneurons in Parkinson's disease and L-DOPA-induced dyskinesia.Neurosci Biobehav Rev. 2018 Sep;92:67-82. doi: 10.1016/j.neubiorev.2018.05.021. Epub 2018 May 18. Neurosci Biobehav Rev. 2018. PMID: 29782883 Review.
Cited by
-
Recurrent Implication of Striatal Cholinergic Interneurons in a Range of Neurodevelopmental, Neurodegenerative, and Neuropsychiatric Disorders.Cells. 2021 Apr 15;10(4):907. doi: 10.3390/cells10040907. Cells. 2021. PMID: 33920757 Free PMC article. Review.
-
Decreased Exosomal Acetylcholinesterase Activity in the Plasma of Patients With Parkinson's Disease.Front Aging Neurosci. 2021 May 28;13:665400. doi: 10.3389/fnagi.2021.665400. eCollection 2021. Front Aging Neurosci. 2021. PMID: 34122043 Free PMC article.
-
Fluorescence microscopy imaging of a neurotransmitter receptor and its cell membrane lipid milieu.Front Mol Biosci. 2022 Nov 28;9:1014659. doi: 10.3389/fmolb.2022.1014659. eCollection 2022. Front Mol Biosci. 2022. PMID: 36518846 Free PMC article. Review.
-
Dopaminergic and Cholinergic Modulation of Large Scale Networks in silico Using Snudda.Front Neural Circuits. 2021 Oct 21;15:748989. doi: 10.3389/fncir.2021.748989. eCollection 2021. Front Neural Circuits. 2021. PMID: 34744638 Free PMC article.
-
A review of the neurotransmitter system associated with cognitive function of the cerebellum in Parkinson's disease.Neural Regen Res. 2024 Feb;19(2):324-330. doi: 10.4103/1673-5374.379042. Neural Regen Res. 2024. PMID: 37488885 Free PMC article. Review.
References
-
- Abercrombie ED, Bonatz AE, and Zigmond MJ (1990). Effects of L-dopa on extracellular dopamine in striatum of normal and 6-hydroxydopamine-treated rats. Brain Res 525, 36–44. - PubMed
-
- Aosaki T, Graybiel AM, and Kimura M (1994). Effect of the nigrostriatal dopamine system on acquired neural responses in the striatum of behaving monkeys. Science 265, 412–415. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
