

TurboID-based proximity labeling reveals that UBR7 is a regulator of N NLR immune receptor-mediated immunity
- PMID: 31324801
- PMCID: PMC6642208
- DOI: 10.1038/s41467-019-11202-z
TurboID-based proximity labeling reveals that UBR7 is a regulator of N NLR immune receptor-mediated immunity
Erratum in
-
Author Correction: TurboID-based proximity labeling reveals that UBR7 is a regulator of N NLR immune receptor-mediated immunity.Nat Commun. 2021 Oct 21;12(1):6200. doi: 10.1038/s41467-021-26441-2. Nat Commun. 2021. PMID: 34675217 Free PMC article. No abstract available.
Abstract
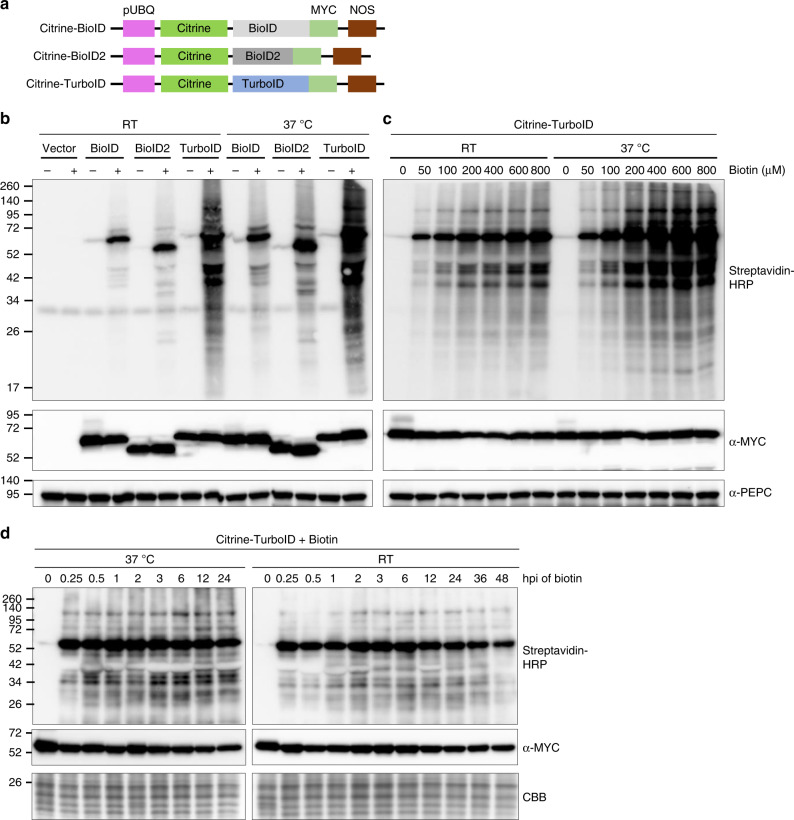
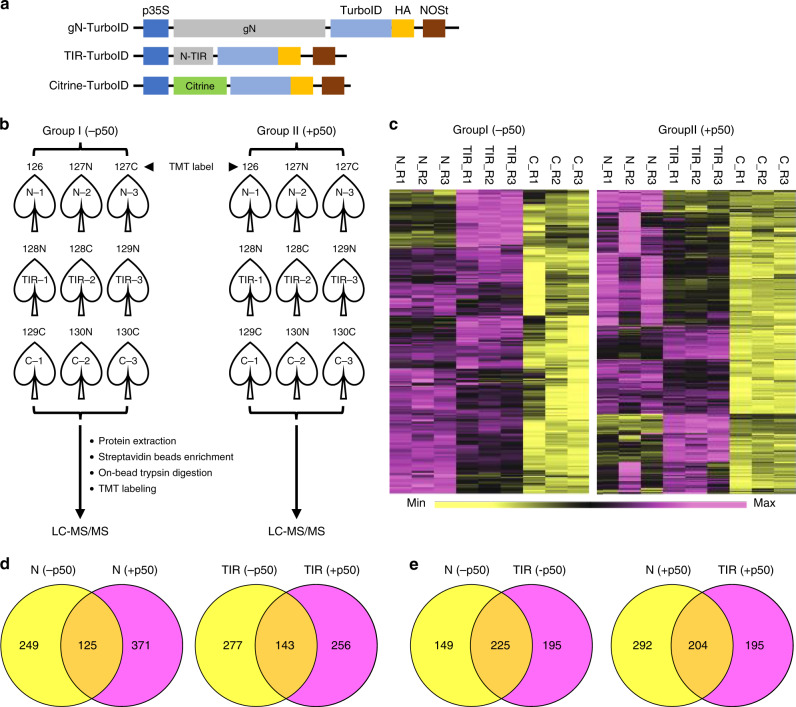
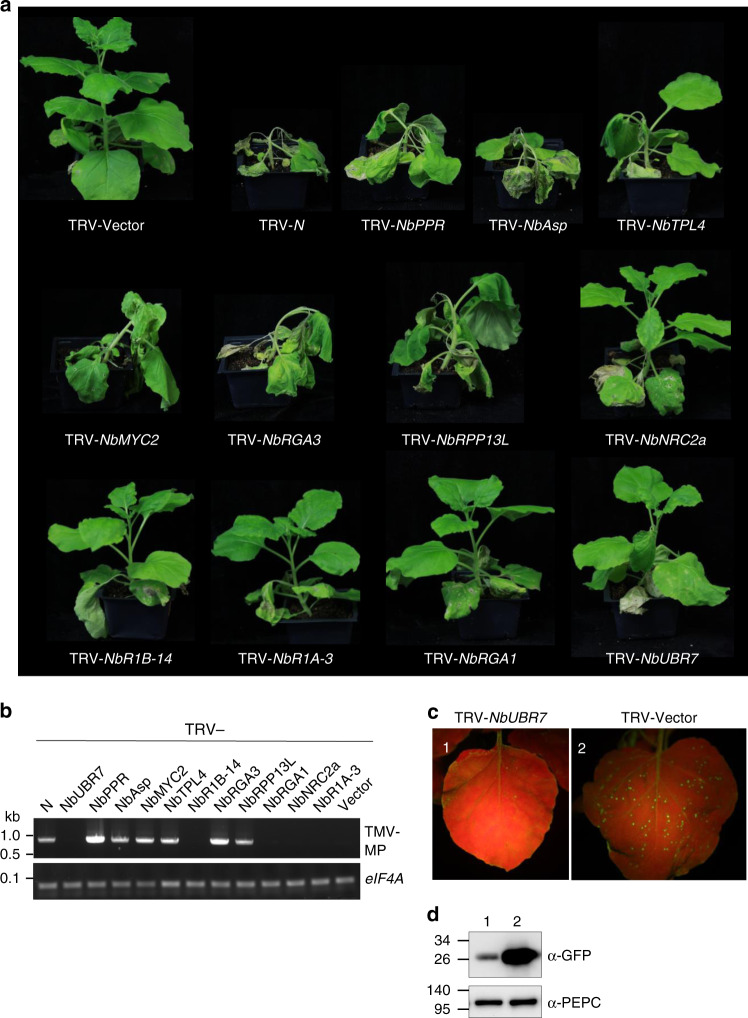
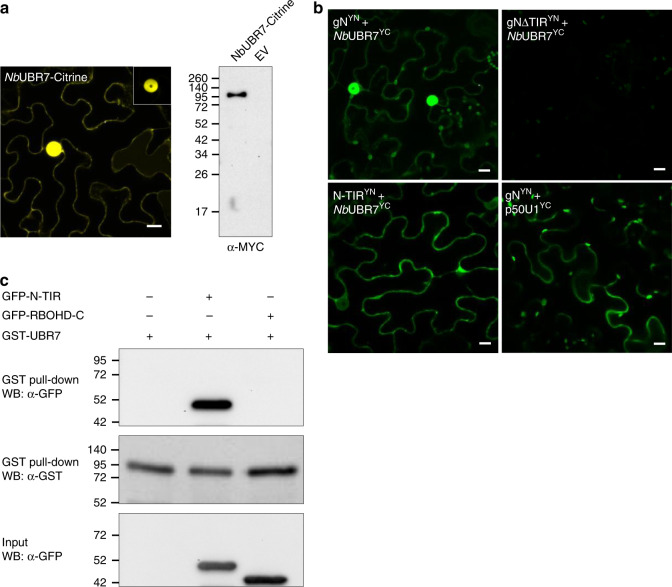
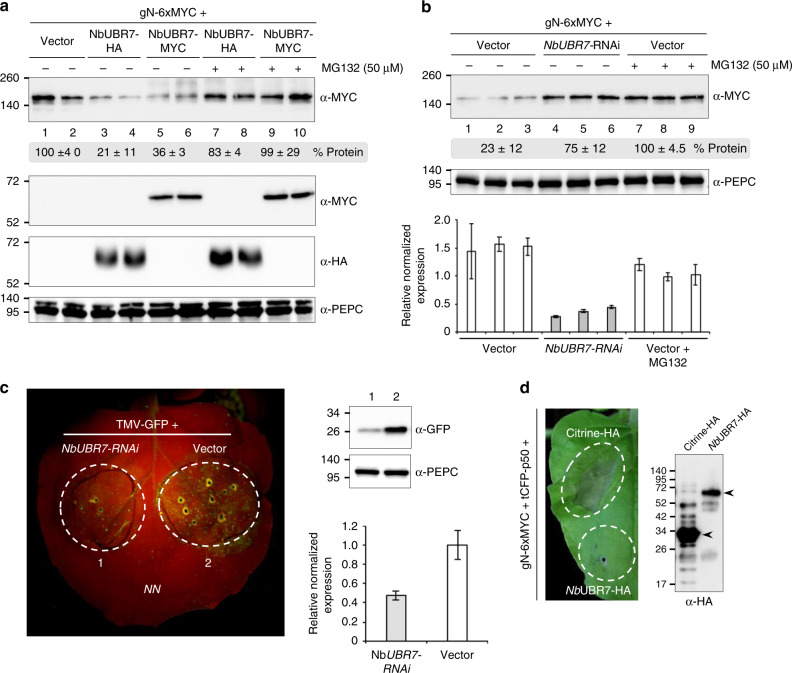
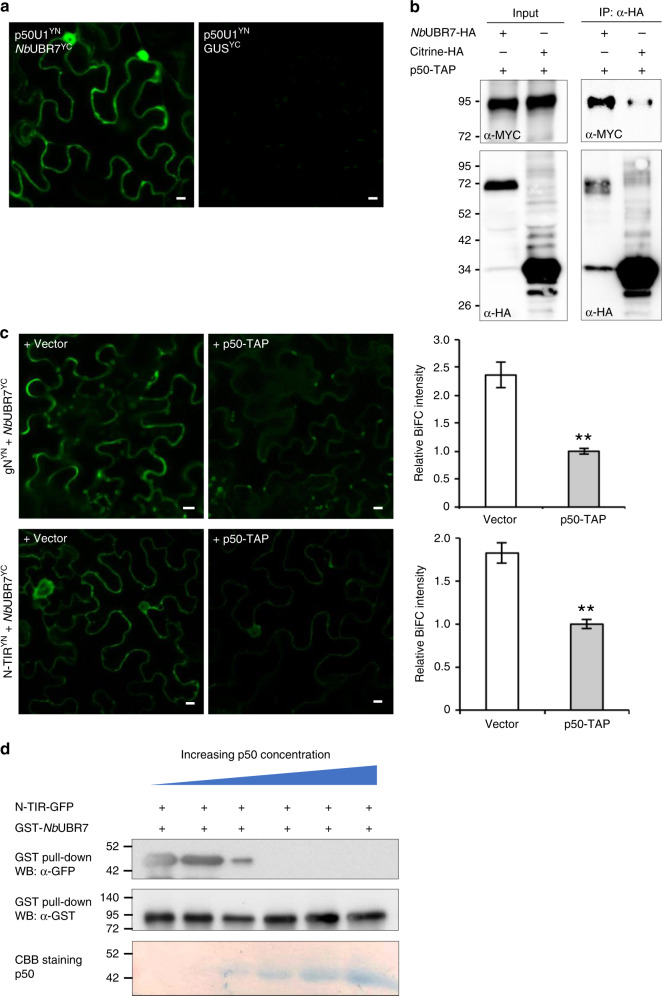
Nucleotide-binding leucine-rich repeat (NLR) immune receptors play a critical role in defence against pathogens in plants and animals. However, we know very little about NLR-interacting proteins and the mechanisms that regulate NLR levels. Here, we used proximity labeling (PL) to identify the proteome proximal to N, which is an NLR that confers resistance to Tobacco mosaic virus (TMV). Evaluation of different PL methods indicated that TurboID-based PL provides more efficient levels of biotinylation than BioID and BioID2 in plants. TurboID-based PL of N followed by quantitative proteomic analysis and genetic screening revealed multiple regulators of N-mediated immunity. Interestingly, a putative E3 ubiquitin ligase, UBR7, directly interacts with the TIR domain of N. UBR7 downregulation leads to an increased amount of N protein and enhanced TMV resistance. TMV-p50 effector disrupts the N-UBR7 interaction and relieves negative regulation of N. These findings demonstrate the utility of TurboID-based PL in plants and the N-interacting proteins we identified enhance our understanding of the mechanisms underlying NLR regulation.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
