

Gains and losses of metabolic function inferred from a phylotranscriptomic analysis of algae
- PMID: 31324835
- PMCID: PMC6642084
- DOI: 10.1038/s41598-019-46869-3
Gains and losses of metabolic function inferred from a phylotranscriptomic analysis of algae
Abstract
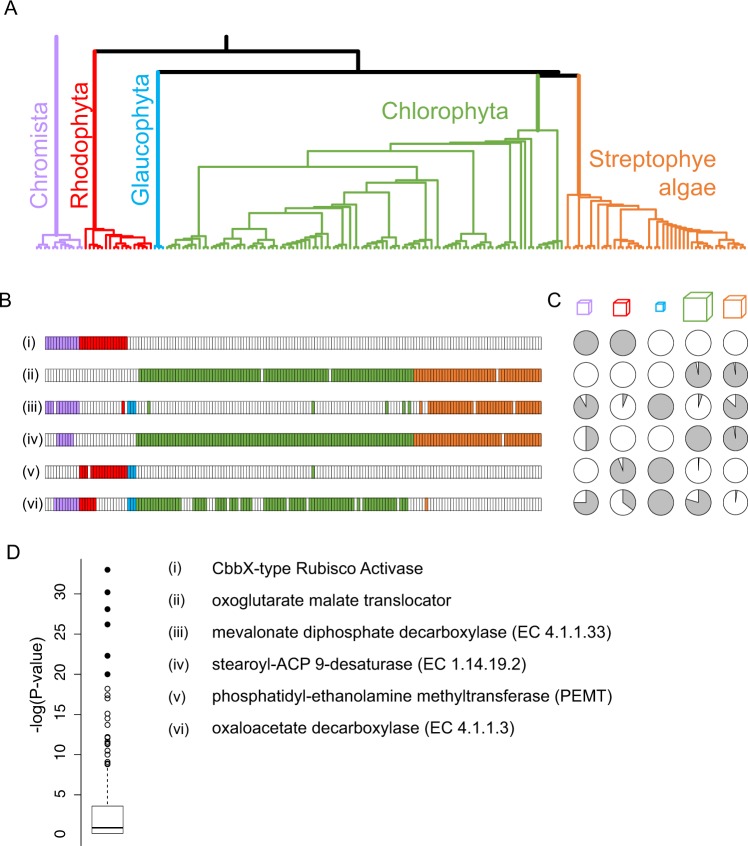
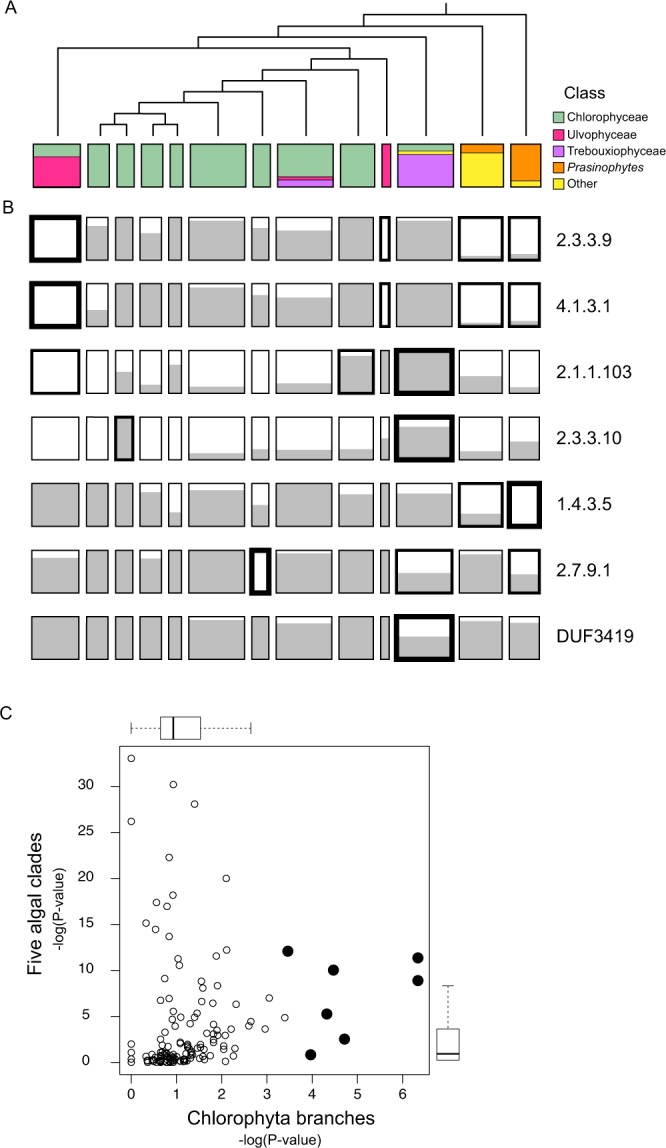
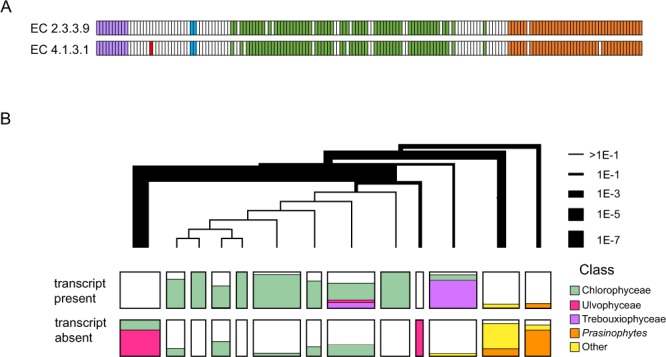
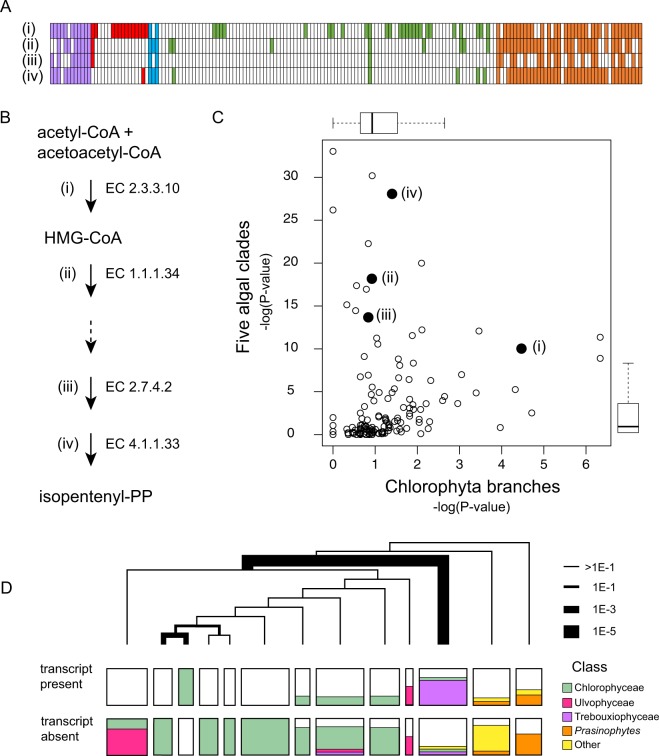
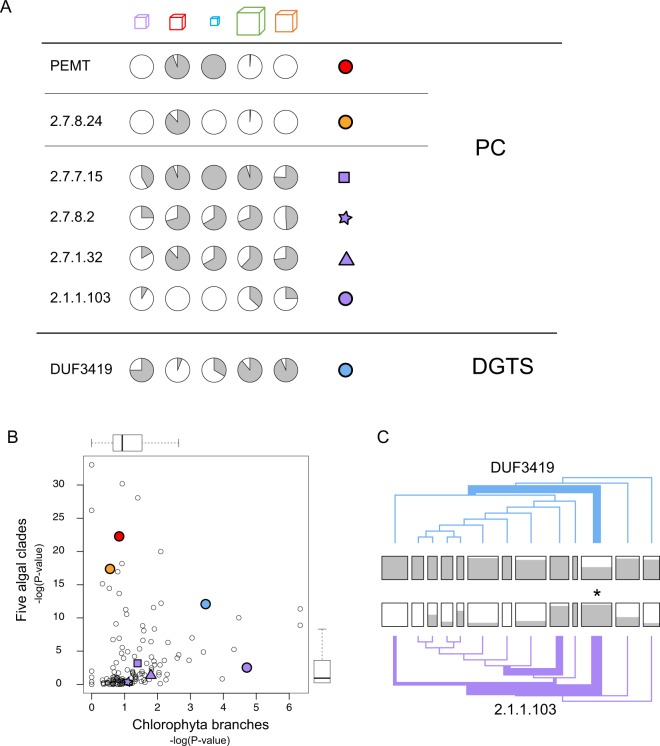
Hidden Markov models representing 167 protein sequence families were used to infer the presence or absence of homologs within the transcriptomes of 183 algal species/strains. Statistical analyses of the distribution of HMM hits across major clades of algae, or at branch points on the phylogenetic tree of 98 chlorophytes, confirmed and extended known cases of metabolic loss and gain, most notably the loss of the mevalonate pathway for terpenoid synthesis in green algae but not, as we show here, in the streptophyte algae. Evidence for novel events was found as well, most remarkably in the recurrent and coordinated gain or loss of enzymes for the glyoxylate shunt. We find, as well, a curious pattern of retention (or re-gain) of HMG-CoA synthase in chlorophytes that have otherwise lost the mevalonate pathway, suggesting a novel, co-opted function for this enzyme in select lineages. Finally, we find striking, phylogenetically linked distributions of coding sequences for three pathways that synthesize the major membrane lipid phosphatidylcholine, and a complementary phylogenetic distribution pattern for the non-phospholipid DGTS (diacyl-glyceryl-trimethylhomoserine). Mass spectrometric analysis of lipids from 25 species was used to validate the inference of DGTS synthesis from sequence data.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Brodie Juliet, Chan Cheong Xin, De Clerck Olivier, Cock J. Mark, Coelho Susana M., Gachon Claire, Grossman Arthur R., Mock Thomas, Raven John A., Smith Alison G., Yoon Hwan Su, Bhattacharya Debashish. The Algal Revolution. Trends in Plant Science. 2017;22(8):726–738. doi: 10.1016/j.tplants.2017.05.005. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
