

A permethrin metabolite is associated with adaptive immune responses in Gulf War Illness
- PMID: 31325531
- PMCID: PMC7155744
- DOI: 10.1016/j.bbi.2019.07.015
A permethrin metabolite is associated with adaptive immune responses in Gulf War Illness
Abstract
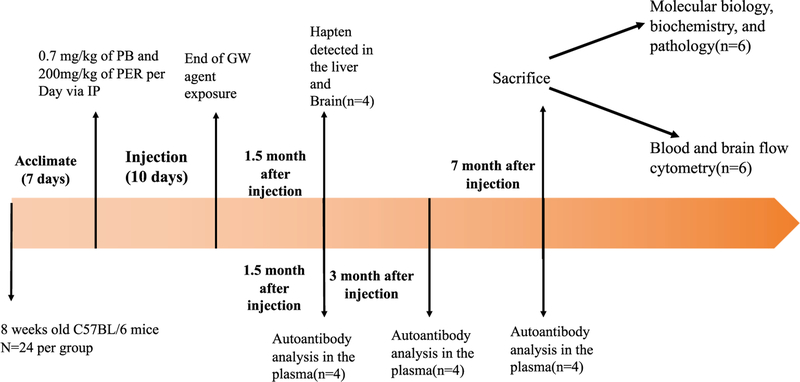
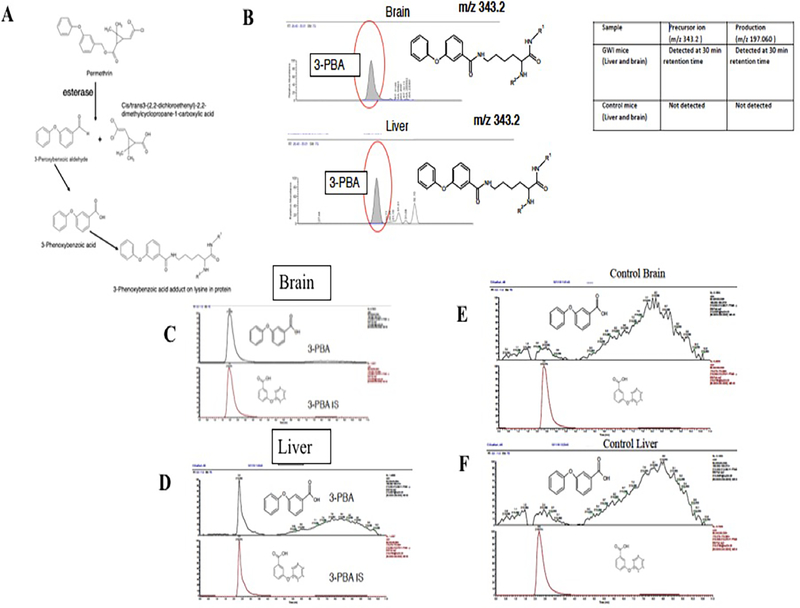
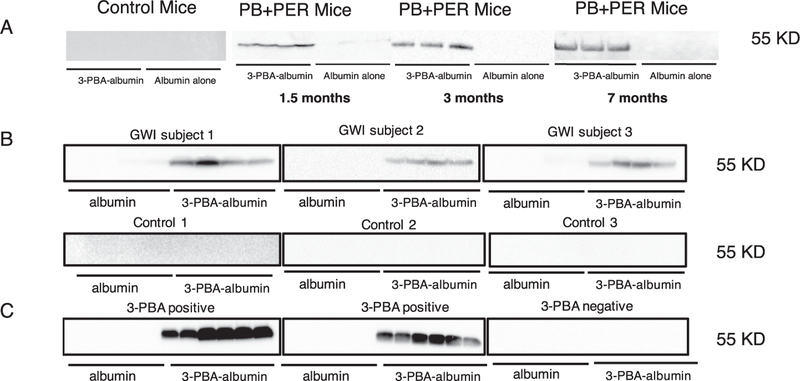
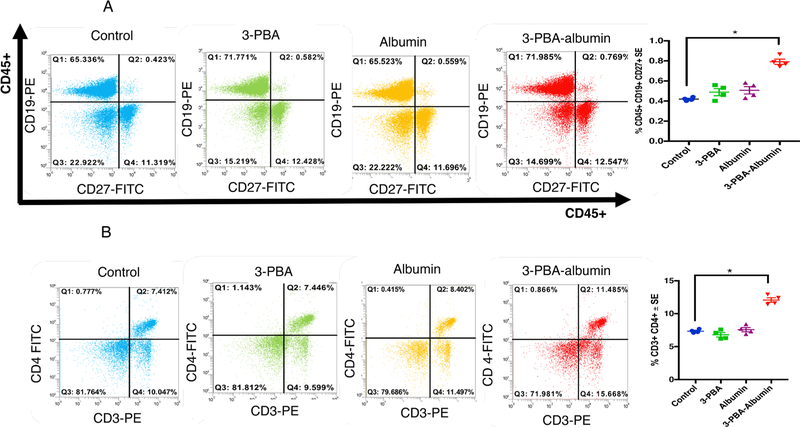
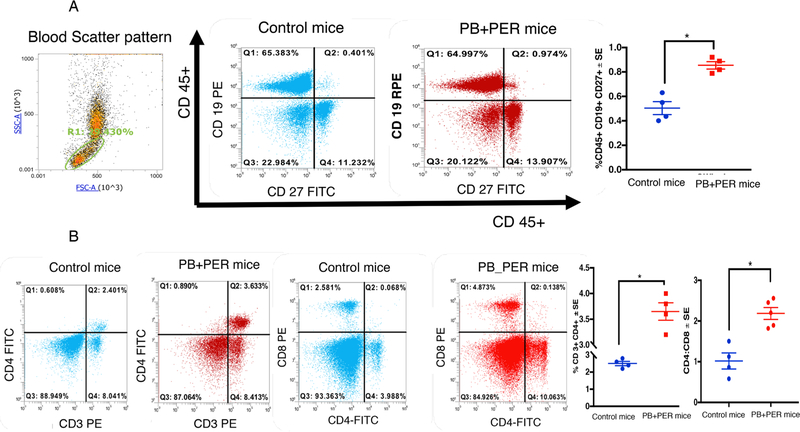
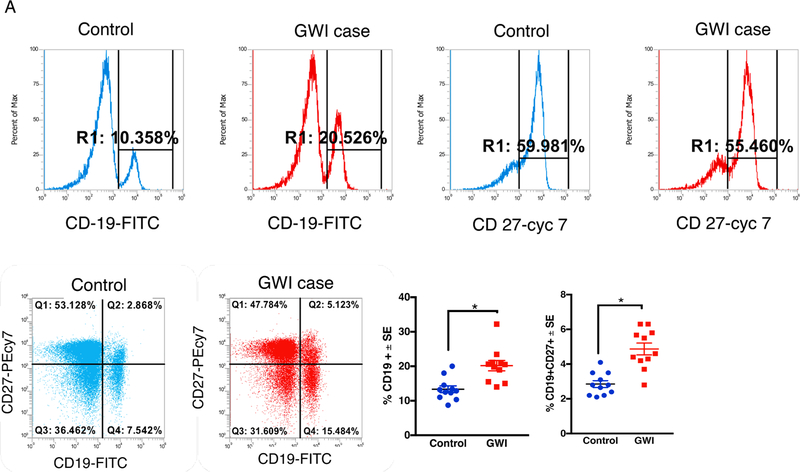
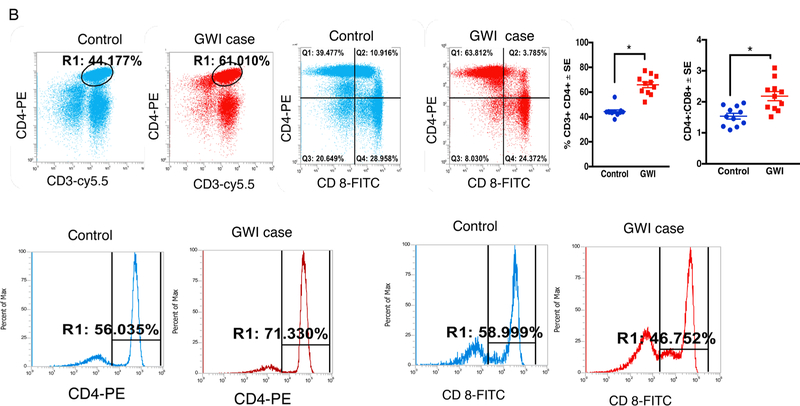
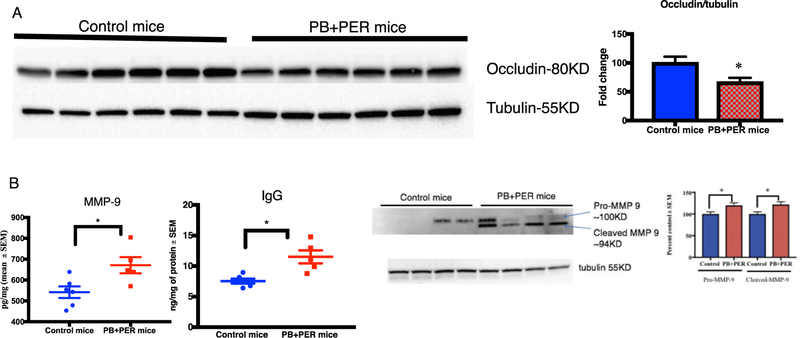
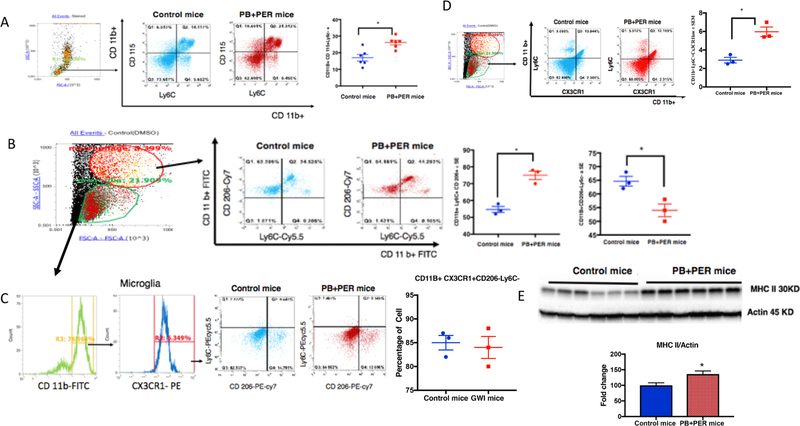
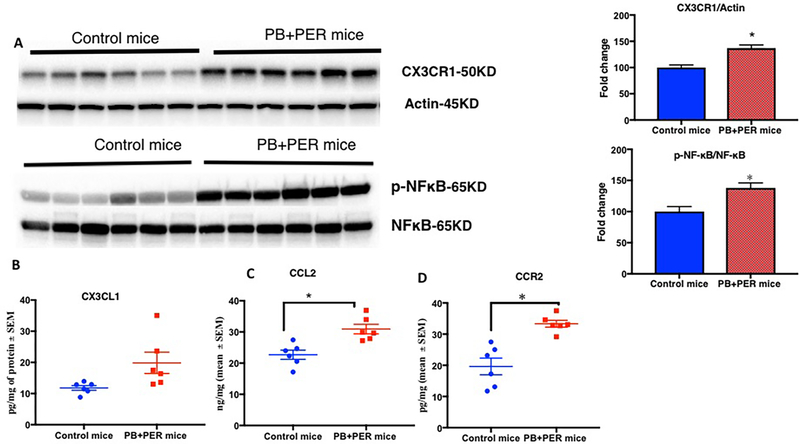
Gulf War Illness (GWI), affecting 30% of veterans from the 1991 Gulf War (GW), is a multi-symptom illness with features similar to those of patients with autoimmune diseases. The objective of the current work is to determine if exposure to GW-related pesticides, such as permethrin (PER), activates peripheral and central nervous system (CNS) adaptive immune responses. In the current study, we focused on a PER metabolite, 3-phenoxybenzoic acid (3-PBA), as this is a common metabolite previously shown to form adducts with endogenous proteins. We observed the presence of 3-PBA and 3-PBA modified lysine of protein peptides in the brain, blood and liver of pyridostigmine bromide (PB) and PER (PB+PER) exposed mice at acute and chronic post-exposure timepoints. We tested whether 3-PBA-haptenated albumin (3-PBA-albumin) can activate immune cells since it is known that chemically haptenated proteins can stimulate immune responses. We detected autoantibodies against 3-PBA-albumin in plasma from PB + PER exposed mice and veterans with GWI at chronic post-exposure timepoints. We also observed that in vitro treatment of blood with 3-PBA-albumin resulted in the activation of B- and T-helper lymphocytes and that these immune cells were also increased in blood of PB + PER exposed mice and veterans with GWI. These immune changes corresponded with elevated levels of infiltrating monocytes in the brain and blood of PB + PER exposed mice which coincided with alterations in the markers of blood-brain barrier disruption, brain macrophages and neuroinflammation. These studies suggest that pesticide exposure associated with GWI may have resulted in the activation of the peripheral and CNS adaptive immune responses, possibly contributing to an autoimmune-type phenotype in veterans with GWI.
Keywords: Autoantibody; Gulf War Illness; Hapten; Permethrin; Pesticide.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest
The author(s) declares no competing interests. The contents do not represent the views of the Department of Veterans Affairs or the United States Government.
Figures
References
-
- Abdel-Rahman A, et al., 2004. Stress and combined exposure to low doses of pyridostigmine bromide, DEET, and permethrin produce neurochemical and neuropathological alterations in cerebral cortex, hippocampus, and cerebellum. J. Toxicol. Environ. Health, Part A 67 (2), 163–192. 10.1080/15287390490264802. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
