

Protein lysine de-2-hydroxyisobutyrylation by CobB in prokaryotes
- PMID: 31328167
- PMCID: PMC6636992
- DOI: 10.1126/sciadv.aaw6703
Protein lysine de-2-hydroxyisobutyrylation by CobB in prokaryotes
Abstract
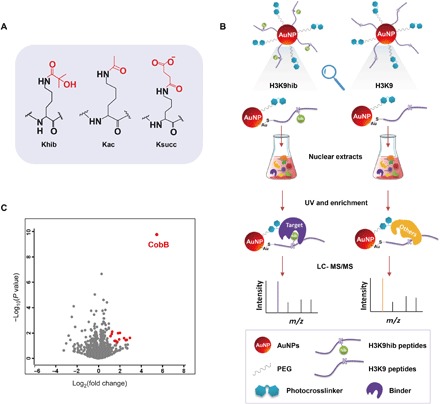
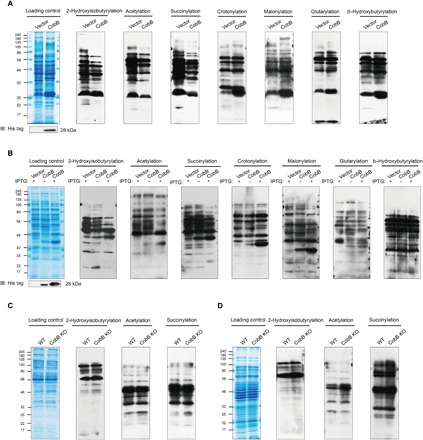
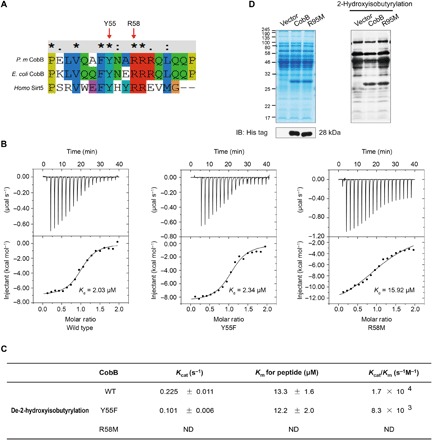
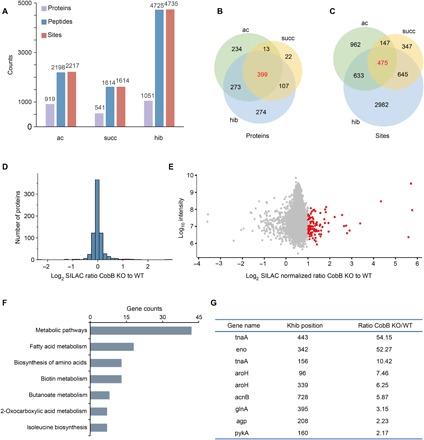
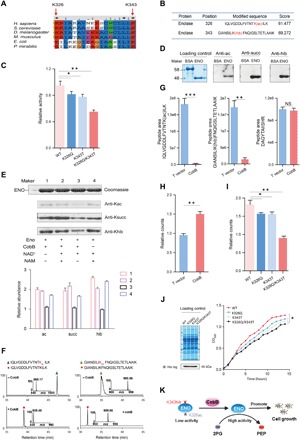
Lysine 2-hydroxyisobutyrylation (Khib) has recently been shown to be an evolutionarily conserved histone mark. Here, we report that CobB serves as a lysine de-2-hydroxyisobutyrylation enzyme that regulates glycolysis and cell growth in prokaryotes. We identified the specific binding of CobB to Khib using a novel self-assembled multivalent photocrosslinking peptide probe and demonstrated that CobB can catalyze lysine de-2-hydroxyisobutyrylation both in vivo and in vitro. R58 of CobB is a critical site for its de-2-hydroxyisobutyrylase activity. Using a quantitative proteomics approach, we identified 99 endogenous substrates that are targeted by CobB for de-2-hydroxyisobutyrylation. We further demonstrated that CobB can regulate the catalytic activities of enolase (ENO) by removing K343hib and K326ac of ENO simultaneously, which account for changes of bacterial growth. In brief, our study dissects a Khib-mediated molecular mechanism that is catalyzed by CobB for the regulation of the activity of metabolic enzymes as well as the cell growth of bacteria.
Figures

References
-
- Dai L., Peng C., Montellier E., Lu Z., Chen Y., Ishii H., Debernardi A., Buchou T., Rousseaux S., Jin F., Sabari B. R., Deng Z., Allis C. D., Ren B., Khochbin S., Zhao Y., Lysine 2-hydroxyisobutyrylation is a widely distributed active histone mark. Nat. Chem. Biol. 10, 365–373 (2014). - PubMed
-
- Montellier E., Rousseaux S., Zhao Y., Khochbin S., Histone crotonylation specifically marks the haploid male germ cell gene expression program. Bioessays 34, 187–193 (2012). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
