

LncRNA MEG3 inhibits cell proliferation and induces apoptosis in laryngeal cancer via miR-23a/APAF-1 axis
- PMID: 31328388
- PMCID: PMC6787452
- DOI: 10.1111/jcmm.14549
LncRNA MEG3 inhibits cell proliferation and induces apoptosis in laryngeal cancer via miR-23a/APAF-1 axis
Abstract
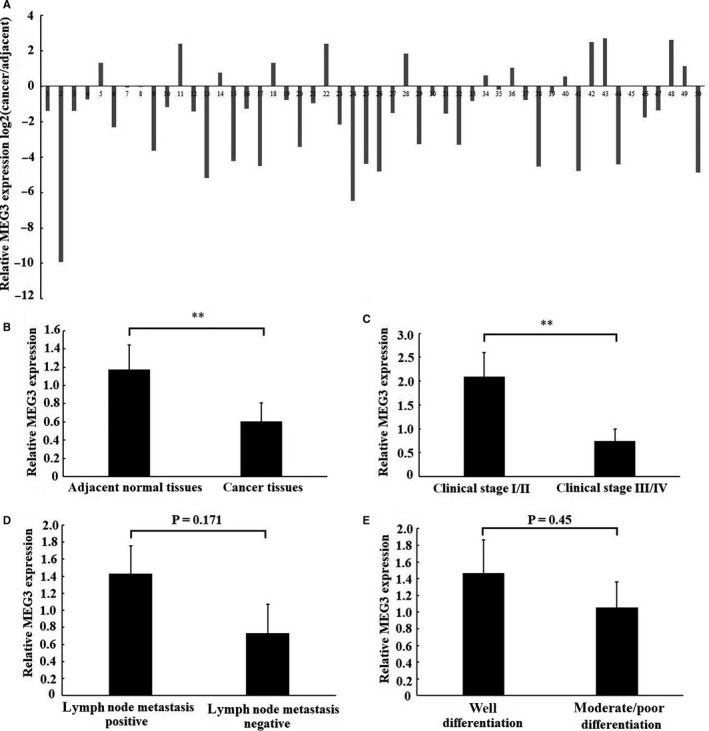
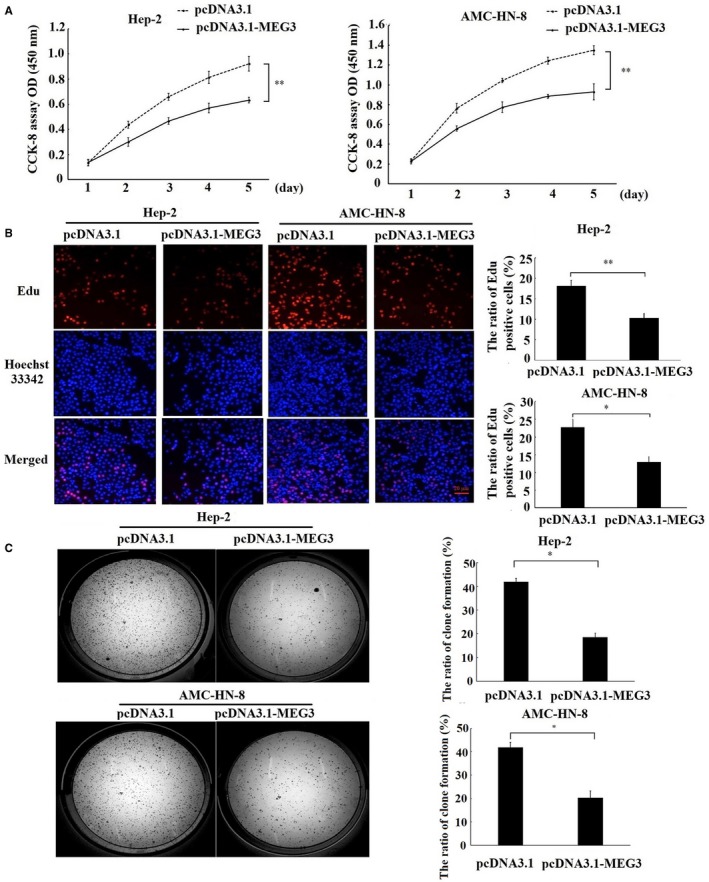
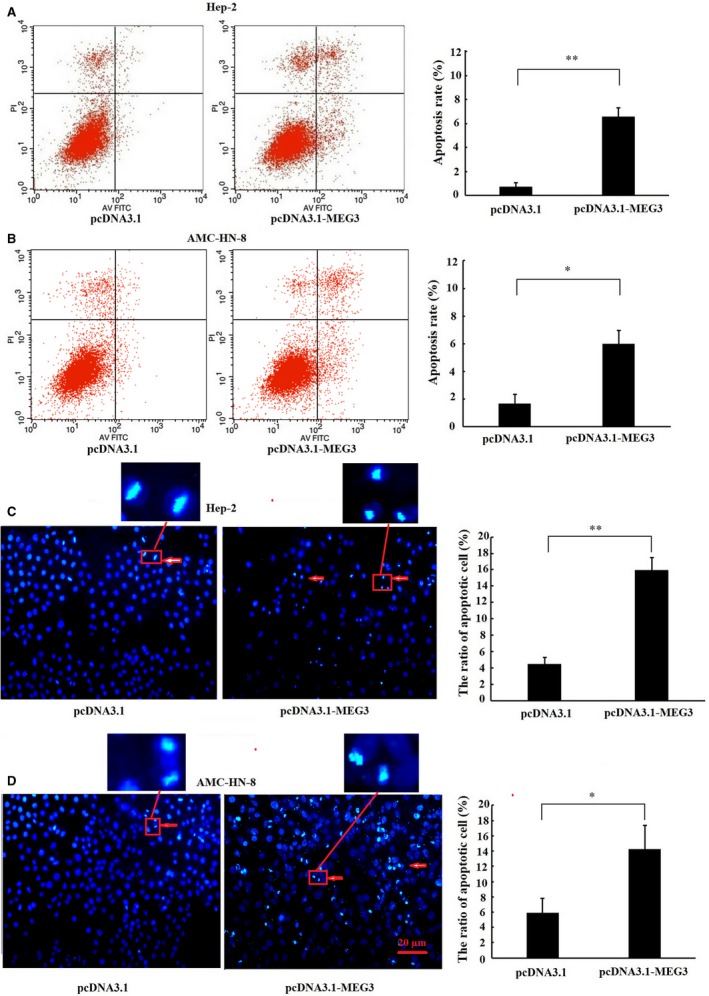
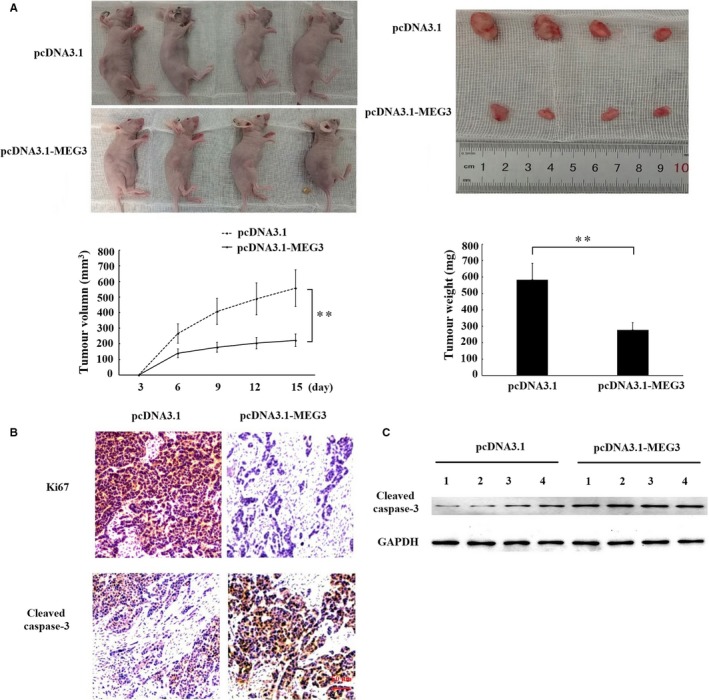
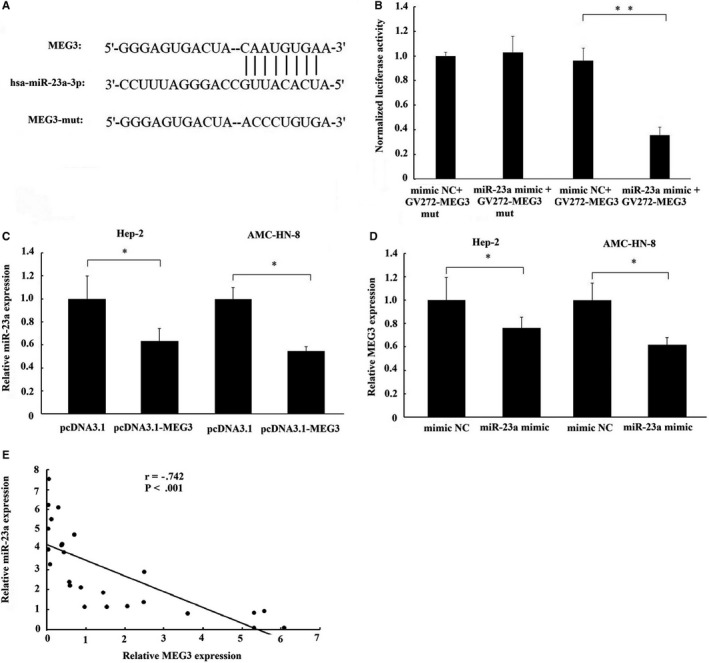
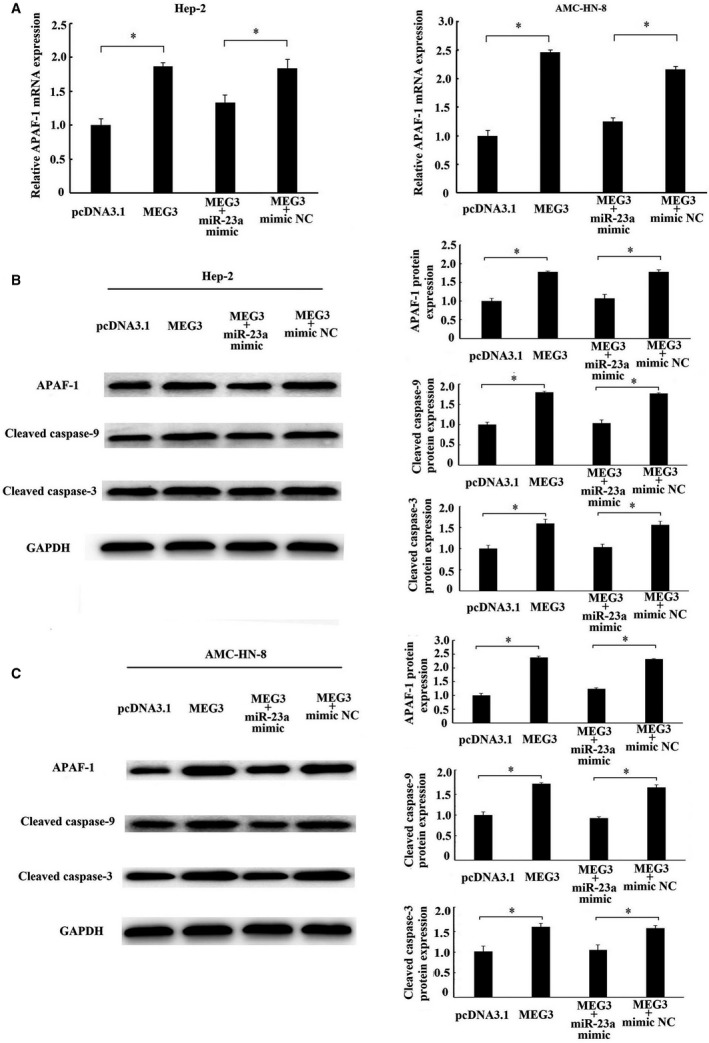
Long non-coding RNA (LncRNA) MEG3 serves a regulatory role in the progression of several types of cancer, but the role of MEG3 in laryngeal cancer is still unknown. The aim of this study was to explore the regulatory role and mechanism of MEG3 in laryngeal cancer. MEG3 expression in 50 laryngeal cancer tissue samples was detected by reverse transcription-quantitative polymerase chain reaction (RT-qPCR). The effects of MEG3 overexpression on laryngeal cancer cells were investigated in vitro and in vivo. The mechanism of competitive endogenous RNA (ceRNA) was validated through luciferase reporter assay, RT-qPCR and Western blotting. MEG3 was down-regulated in laryngeal cancer tissues, and the low MEG3 expression was associated with advanced clinical stage. Additionally, MEG3 overexpression inhibited the proliferation and induced the apoptosis of laryngeal cancer cells in vitro and in vivo. Particularly, MEG3 bound to miR-23a specifically and a reciprocal negative regulation existed between miR-23a and MEG3. Moreover, MEG3 up-regulated apoptotic protease activating factor-1 (APAF-1), a known miR-23a's target, thereby leading to the activation of caspase-9 and caspase-3. Meanwhile, these activated effects were rescued by miR-23a overexpression. In conclusion, the present study demonstrated that MEG3 functions as a novel tumour suppressive LncRNA in laryngeal cancer for the first time. Furthermore, MEG3 may act as a ceRNA to regulate APAF-1 expression by competitive binding to miR-23a, thereby regulating the progression of laryngeal cancer.
Keywords: apoptotic protease activating factor-1; competitive endogenous RNA; laryngeal cancer; maternally expressed gene 3; miRNA-23a.
© 2019 The Authors. Journal of Cellular and Molecular Medicine published by John Wiley & Sons Ltd and Foundation for Cellular and Molecular Medicine.
Conflict of interest statement
The authors have no conflict of interest.
Figures
References
-
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016;66:7‐30. - PubMed
-
- Garavello W, Turati F, Bosetti C, et al. Family history of cancer and the risk of laryngeal cancer: a case‐control study from Italy and Switzerland. Int J Cancer. 2012;130:665‐670. - PubMed
-
- Todorova TA, Jordanov SH, Stancheva GS, et al. Mutational status of CDKN2A and TP53 genes in laryngeal squamous cell carcinoma. Pathol Oncol Res. 2015;21:413‐421. - PubMed
-
- Fanjul‐Fernández M, Quesada V, Cabanillas R, et al. Cell‐cell adhesion genes CTNNA2 and CTNNA3 are tumour suppressors frequently mutated in laryngeal carcinomas. Nat Commun. 2013;4:2531. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
