

Regulation of phosphoribosyl ubiquitination by a calmodulin-dependent glutamylase
- PMID: 31330531
- PMCID: PMC6855250
- DOI: 10.1038/s41586-019-1439-1
Regulation of phosphoribosyl ubiquitination by a calmodulin-dependent glutamylase
Abstract
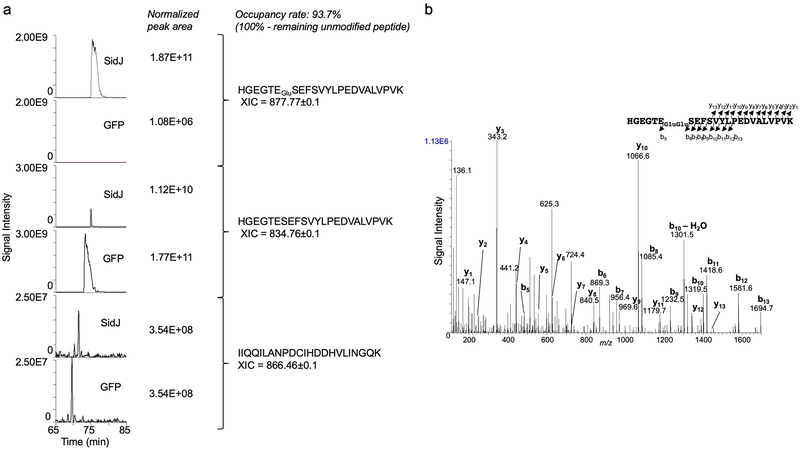
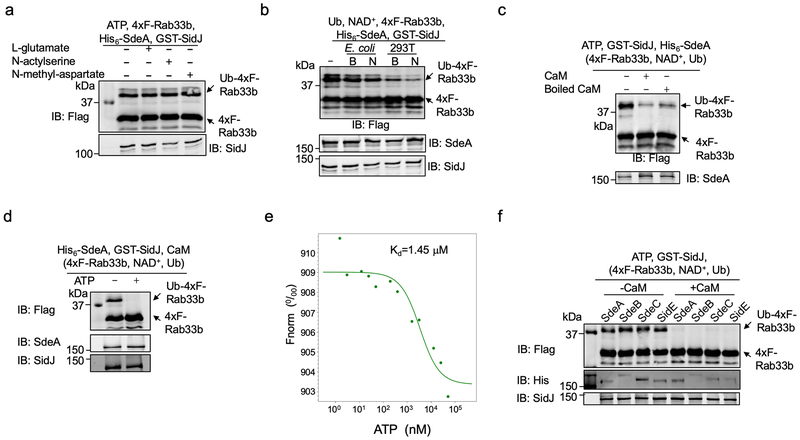
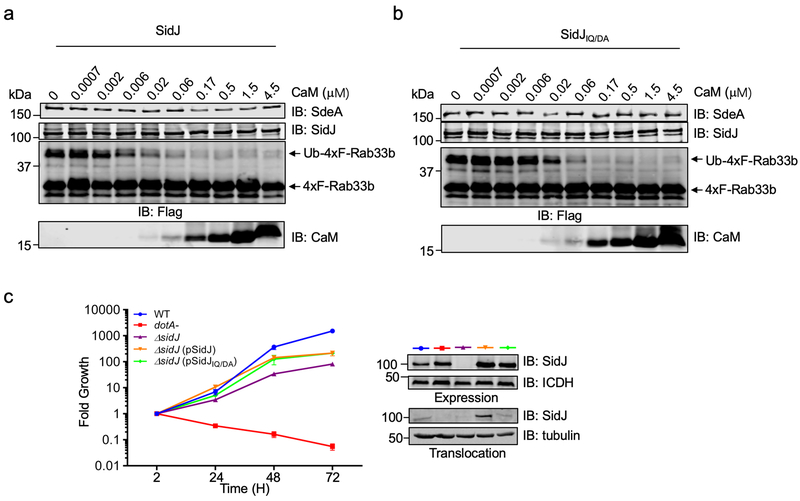
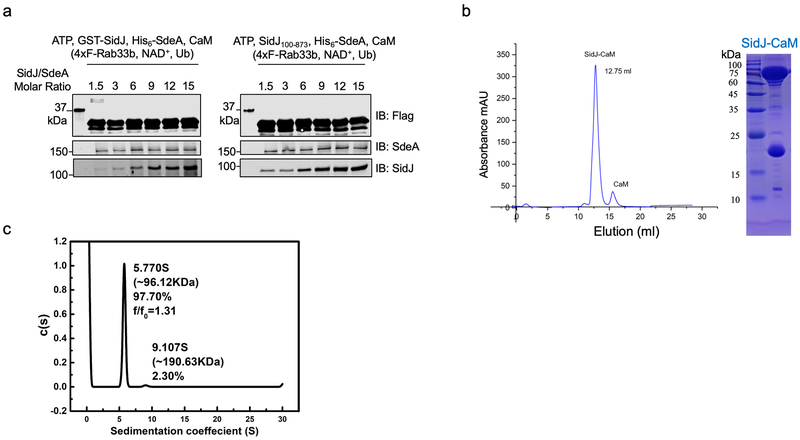
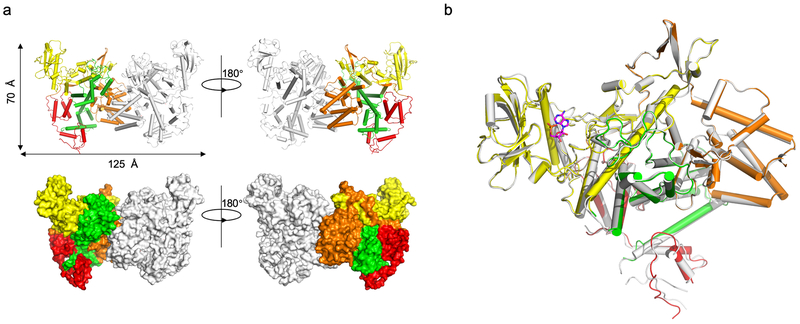
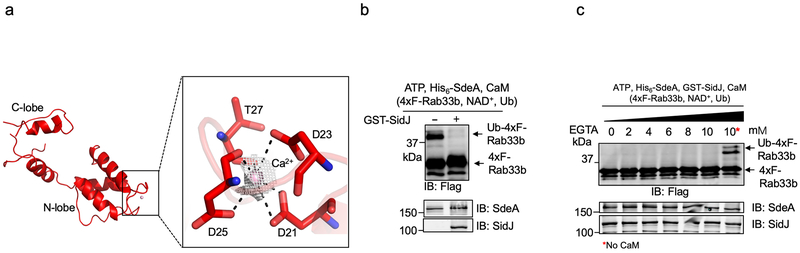
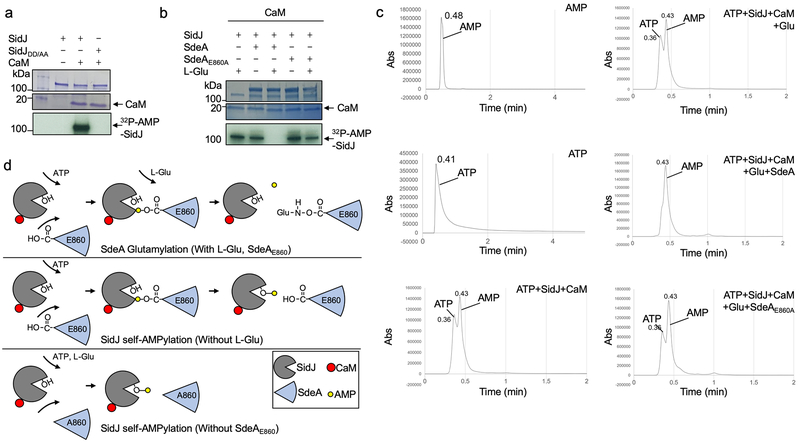
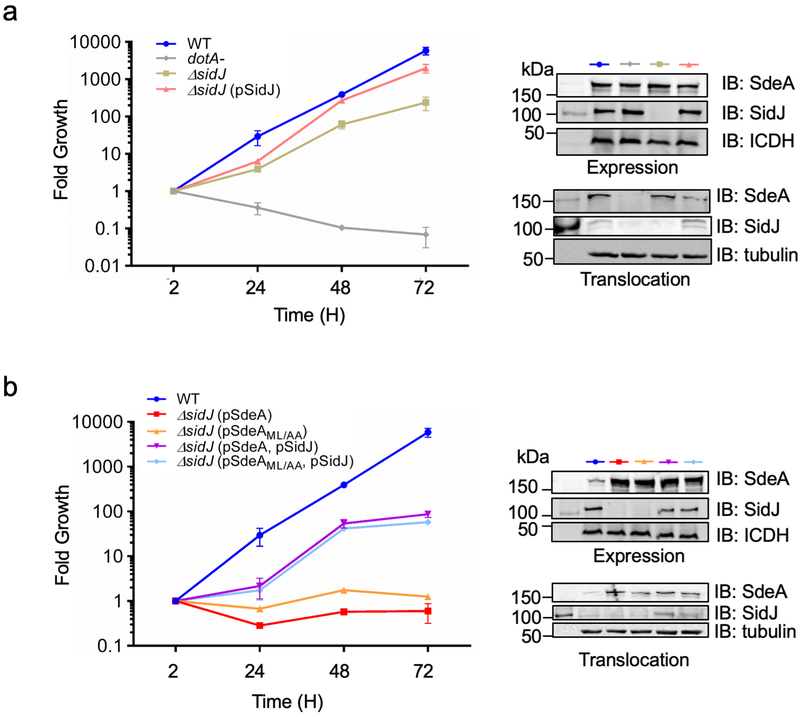
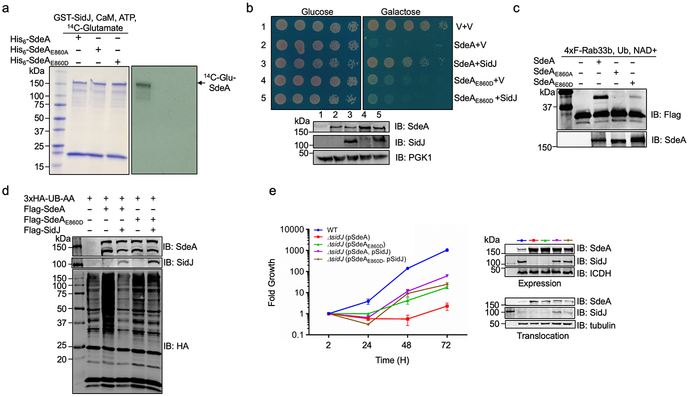
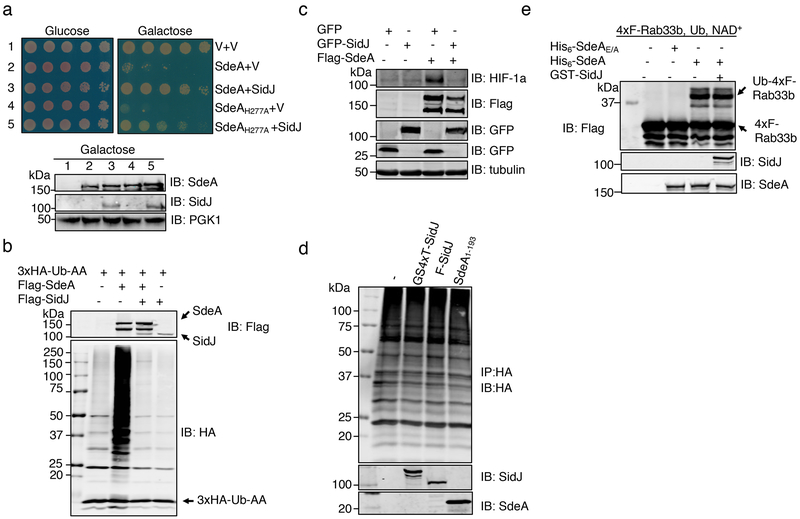
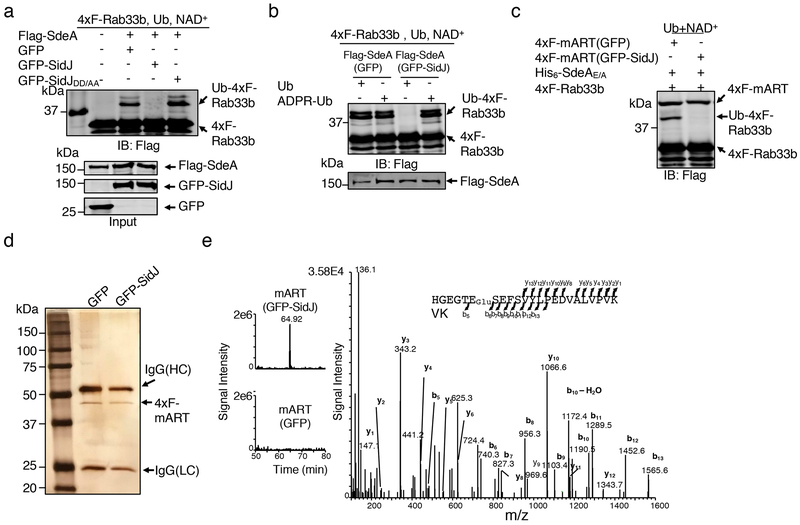
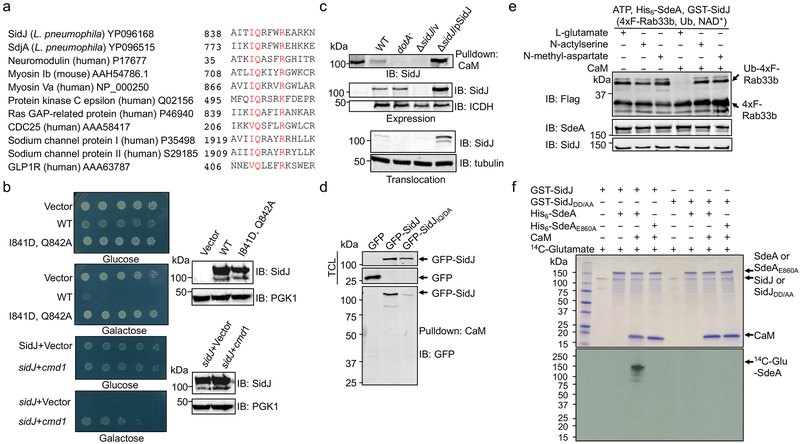
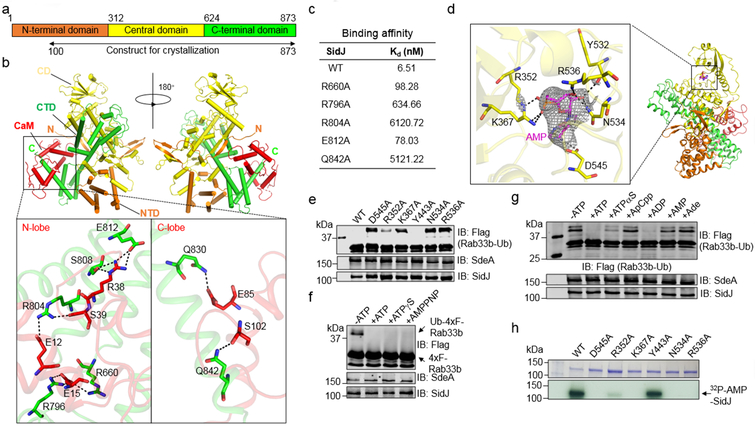
The bacterial pathogen Legionella pneumophila creates an intracellular niche permissive for its replication by extensively modulating host-cell functions using hundreds of effector proteins delivered by its Dot/Icm secretion system1. Among these, members of the SidE family (SidEs) regulate several cellular processes through a unique phosphoribosyl ubiquitination mechanism that bypasses the canonical ubiquitination machinery2-4. The activity of SidEs is regulated by another Dot/Icm effector known as SidJ5; however, the mechanism of this regulation is not completely understood6,7. Here we demonstrate that SidJ inhibits the activity of SidEs by inducing the covalent attachment of glutamate moieties to SdeA-a member of the SidE family-at E860, one of the catalytic residues that is required for the mono-ADP-ribosyltransferase activity involved in ubiquitin activation2. This inhibition by SidJ is spatially restricted in host cells because its activity requires the eukaryote-specific protein calmodulin (CaM). We solved a structure of SidJ-CaM in complex with AMP and found that the ATP used in this reaction is cleaved at the α-phosphate position by SidJ, which-in the absence of glutamate or modifiable SdeA-undergoes self-AMPylation. Our results reveal a mechanism of regulation in bacterial pathogenicity in which a glutamylation reaction that inhibits the activity of virulence factors is activated by host-factor-dependent acyl-adenylation.
Figures
Comment in
-
Glutamylation of Bacterial Ubiquitin Ligases by a Legionella Pseudokinase.Trends Microbiol. 2019 Dec;27(12):967-969. doi: 10.1016/j.tim.2019.09.001. Epub 2019 Oct 14. Trends Microbiol. 2019. PMID: 31624006 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
