

Non-canonical DNA/RNA structures during Transcription-Coupled Double-Strand Break Repair: Roadblocks or Bona fide repair intermediates?
- PMID: 31331819
- PMCID: PMC6764918
- DOI: 10.1016/j.dnarep.2019.102661
Non-canonical DNA/RNA structures during Transcription-Coupled Double-Strand Break Repair: Roadblocks or Bona fide repair intermediates?
Abstract
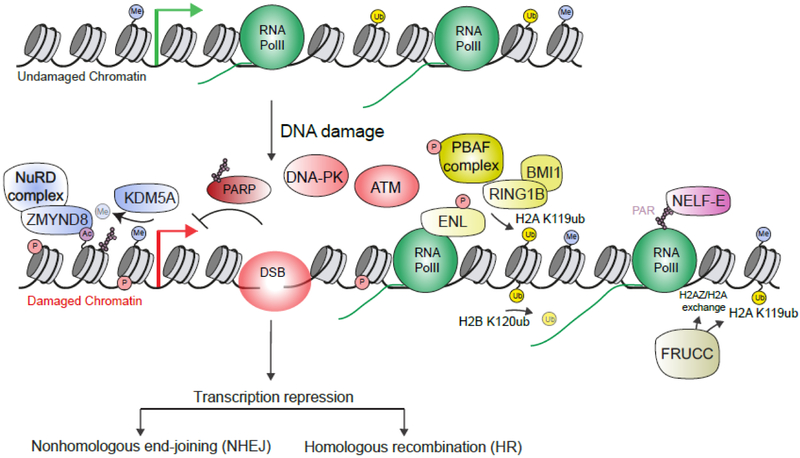
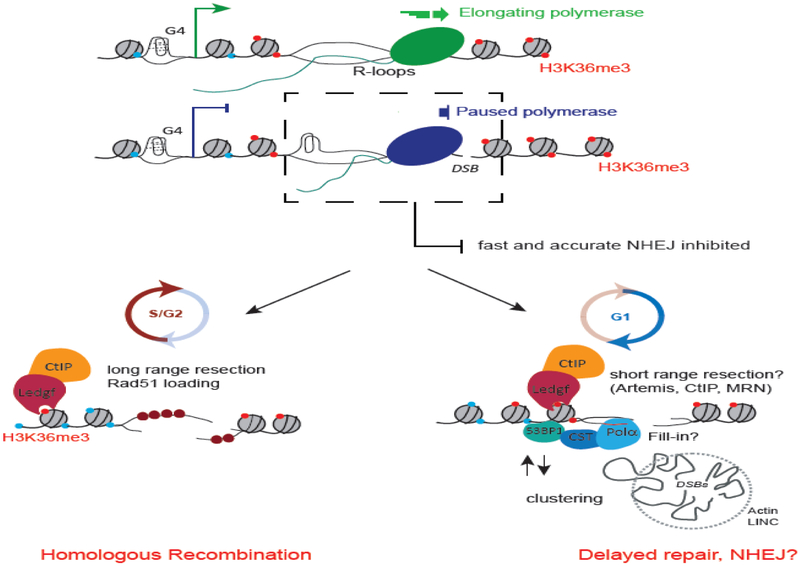
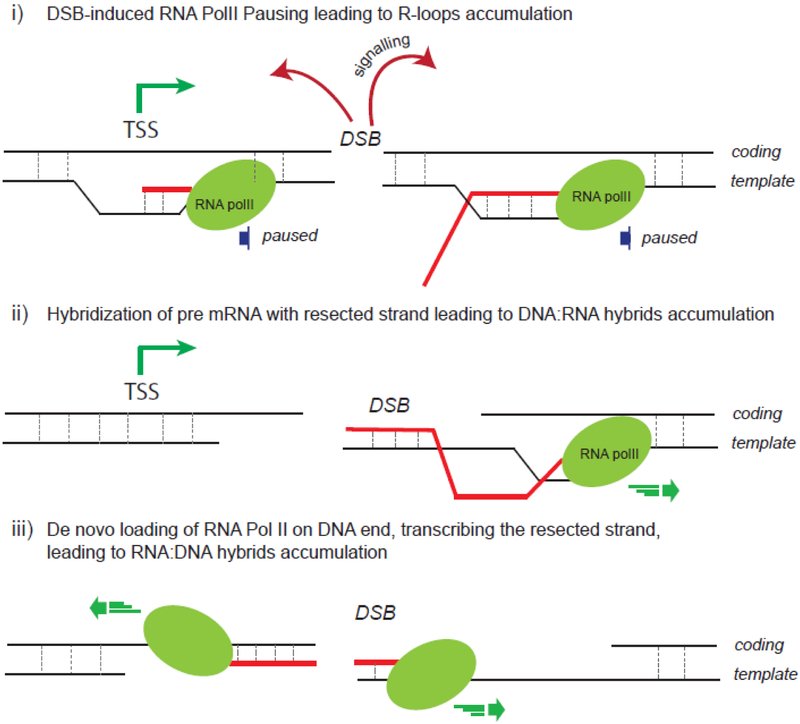
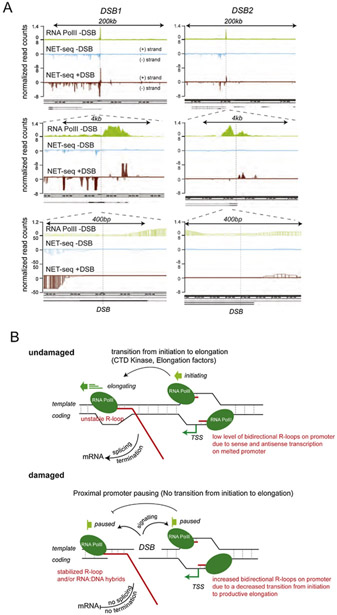
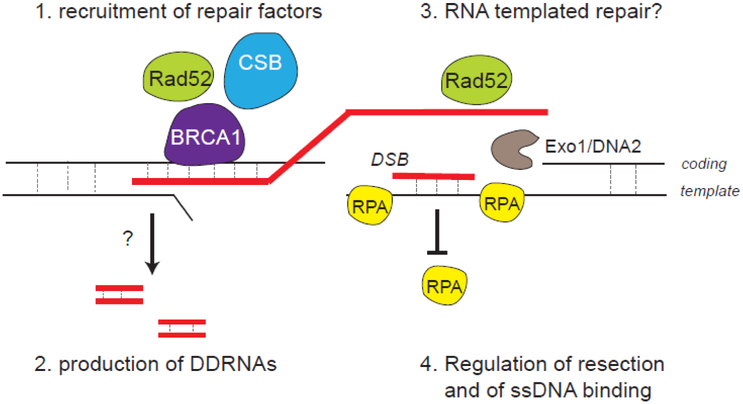
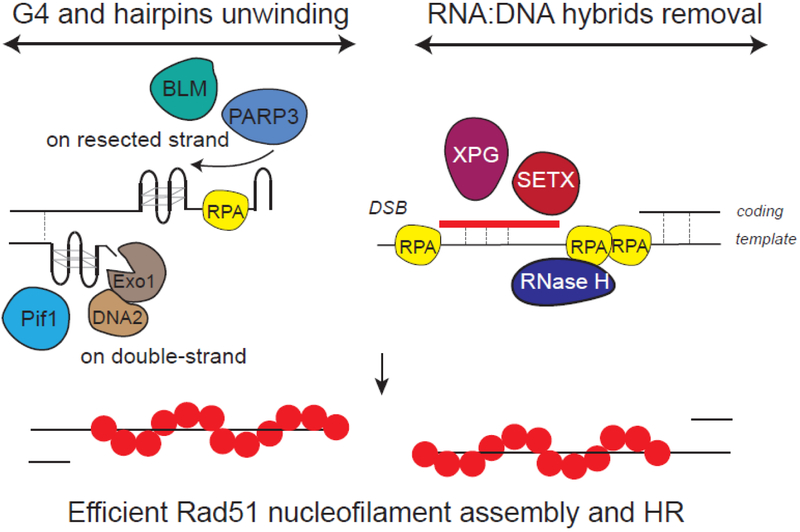
Although long overlooked, it is now well understood that DNA does not systematically assemble into a canonical double helix, known as B-DNA, throughout the entire genome but can also accommodate other structures including DNA hairpins, G-quadruplexes and RNA:DNA hybrids. Notably, these non-canonical DNA structures form preferentially at transcriptionally active loci. Acting as replication roadblocks and being targeted by multiple machineries, these structures weaken the genome and render it prone to damage, including DNA double-strand breaks (DSB). In addition, secondary structures also further accumulate upon DSB formation. Here we discuss the potential functions of pre-existing or de novo formed nucleic acid structures, as bona fide repair intermediates or repair roadblocks, especially during Transcription-Coupled DNA Double-Strand Break repair (TC-DSBR), and provide an update on the specialized protein complexes displaying the ability to remove these structures to safeguard genome integrity.
Keywords: Chromatin; DNA double-strand break repair; G-quadruplex; R-loop; RNA:DNA hybrid; Transcription.
Copyright © 2019 Elsevier B.V. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
