

Elevated faecal 12,13-diHOME concentration in neonates at high risk for asthma is produced by gut bacteria and impedes immune tolerance
- PMID: 31332384
- PMCID: PMC6830510
- DOI: 10.1038/s41564-019-0498-2
Elevated faecal 12,13-diHOME concentration in neonates at high risk for asthma is produced by gut bacteria and impedes immune tolerance
Erratum in
-
Author Correction: Elevated faecal 12,13-diHOME concentration in neonates at high risk for asthma is produced by gut bacteria and impedes immune tolerance.Nat Microbiol. 2019 Nov;4(11):2020. doi: 10.1038/s41564-019-0574-7. Nat Microbiol. 2019. PMID: 31492948
Abstract
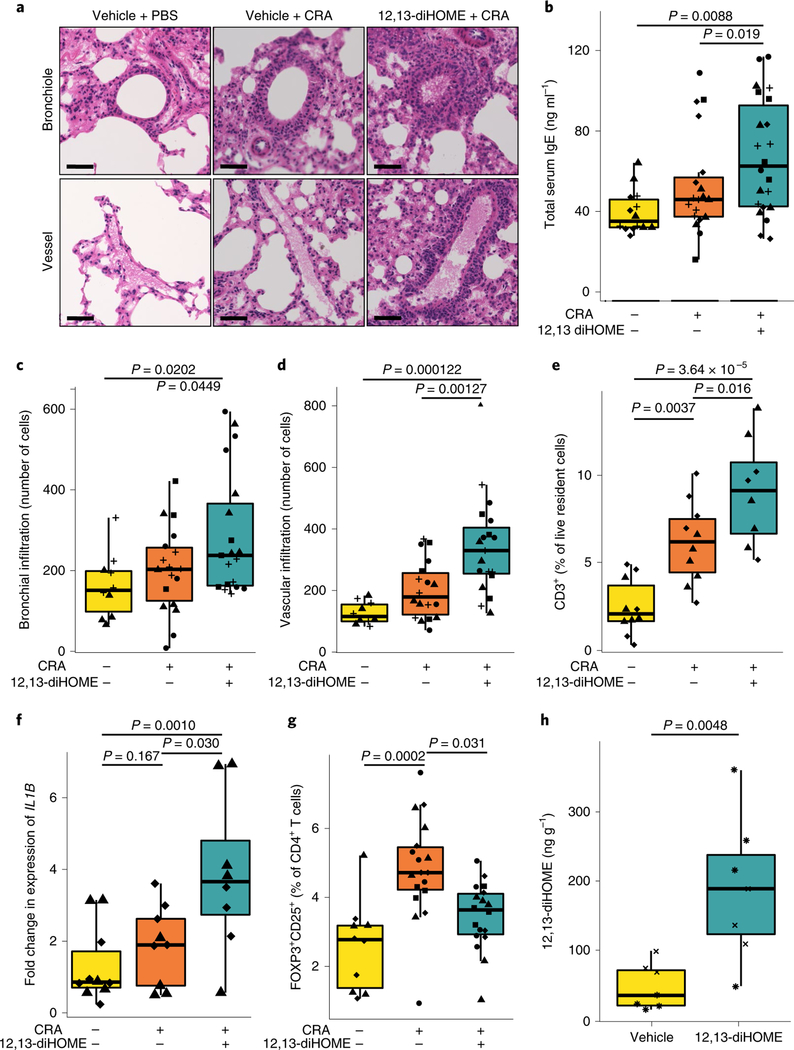
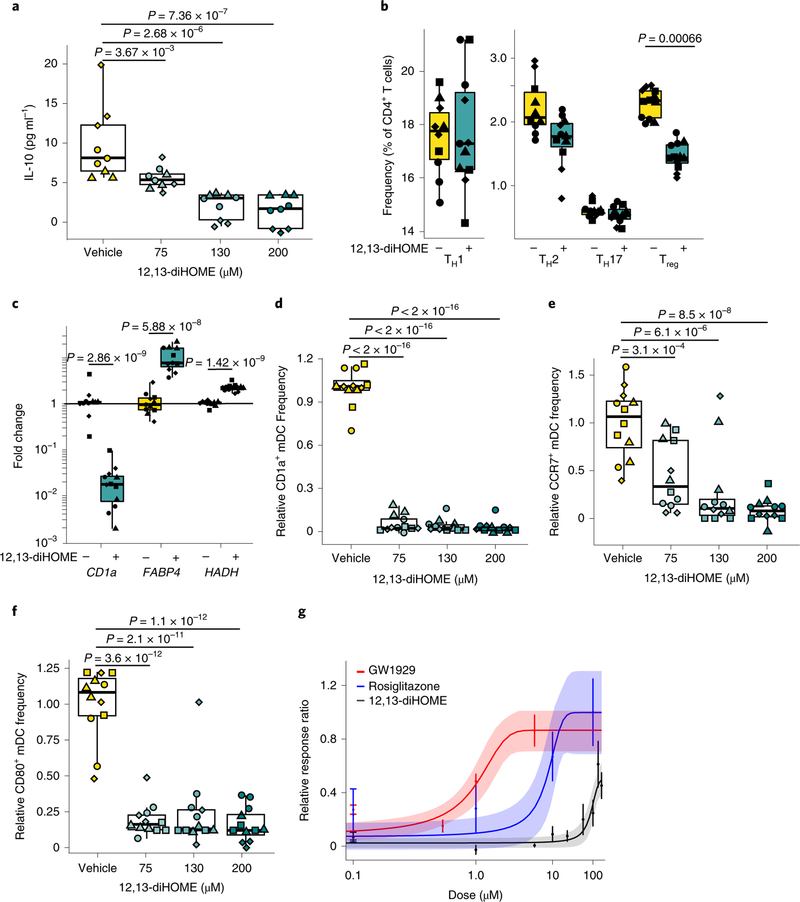
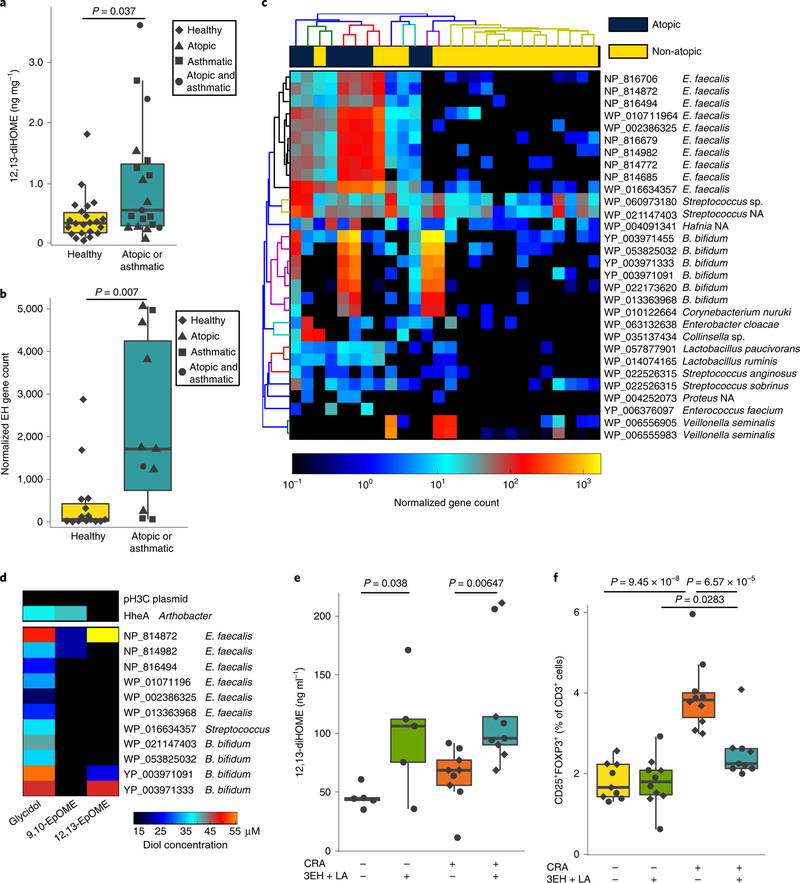
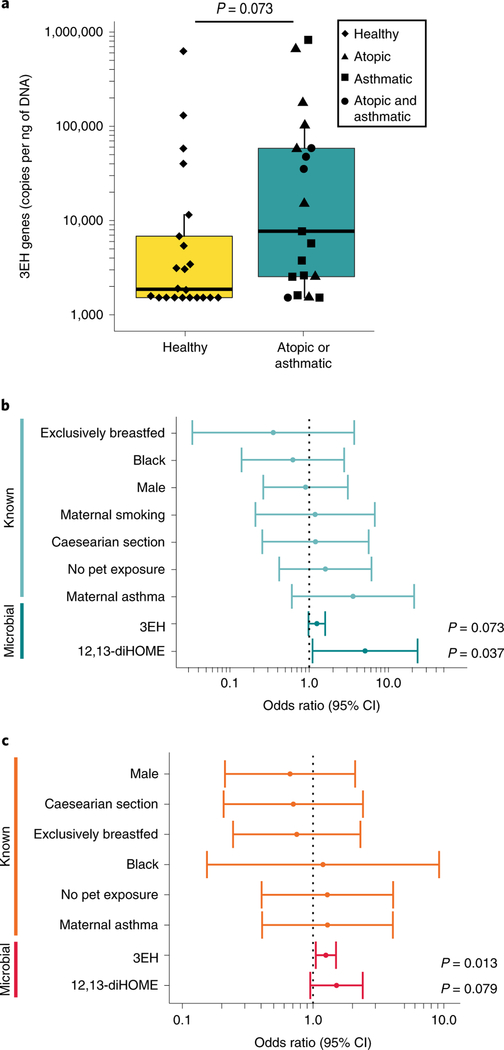
Neonates at risk of childhood atopy and asthma exhibit perturbation of the gut microbiome, metabolic dysfunction and increased concentrations of 12,13-diHOME in their faeces. However, the mechanism, source and contribution of this lipid to allergic inflammation remain unknown. Here, we show that intra-abdominal treatment of mice with 12,13-diHOME increased pulmonary inflammation and decreased the number of regulatory T (Treg) cells in the lungs. Treatment of human dendritic cells with 12,13-diHOME altered expression of PPARγ-regulated genes and reduced anti-inflammatory cytokine secretion and the number of Treg cells in vitro. Shotgun metagenomic sequencing of neonatal faeces indicated that bacterial epoxide hydrolase (EH) genes are more abundant in the gut microbiome of neonates who develop atopy and/or asthma during childhood. Three of these bacterial EH genes (3EH) specifically produce 12,13-diHOME, and treatment of mice with bacterial strains expressing 3EH caused a decrease in the number of lung Treg cells in an allergen challenge model. In two small birth cohorts, an increase in the copy number of 3EH or the concentration of 12,13-diHOME in the faeces of neonates was found to be associated with an increased probability of developing atopy, eczema or asthma during childhood. Our data indicate that elevated 12,13-diHOME concentrations impede immune tolerance and may be produced by bacterial EHs in the neonatal gut, offering a mechanistic link between perturbation of the gut microbiome during early life and atopy and asthma during childhood.
Figures
Comment in
-
Bacterial metabolites shape neonatal immune system.Nat Rev Immunol. 2019 Sep;19(9):537. doi: 10.1038/s41577-019-0207-7. Nat Rev Immunol. 2019. PMID: 31371823 No abstract available.
-
Homing in on 12,13-diHOME in asthma.Nat Microbiol. 2019 Nov;4(11):1774-1775. doi: 10.1038/s41564-019-0599-y. Nat Microbiol. 2019. PMID: 31649356 No abstract available.
References
-
- Yamamoto-Hanada K., Yang L., Narita M., Saito H. & Ohya Y. Influence of antibiotic use in early childhood on asthma and allergic diseases at age 5. Ann. Allergy Asthma Immunol 119, 54–58 (2017). - PubMed
-
- Silvers KM et al. Breastfeeding protects against current asthma up to 6 years of age. J. Pediatr 160, 991–996 (2012). - PubMed
-
- Fall T. et al. Early exposure to dogs and farm animals and the risk of childhood asthma. JAMA Pediatr 169, e153219 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
