

The structure of the influenza A virus genome
- PMID: 31332385
- PMCID: PMC7191640
- DOI: 10.1038/s41564-019-0513-7
The structure of the influenza A virus genome
Abstract
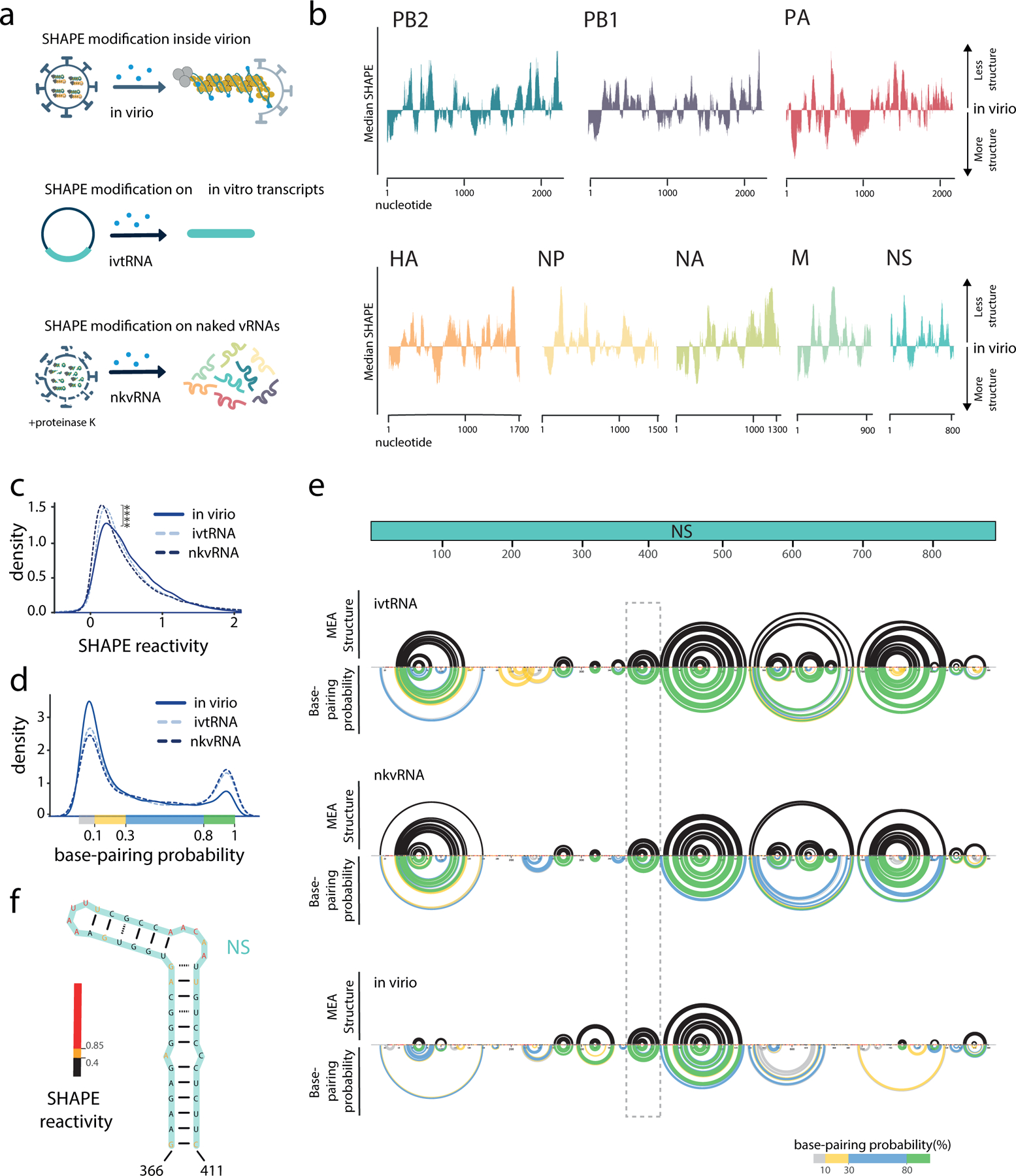
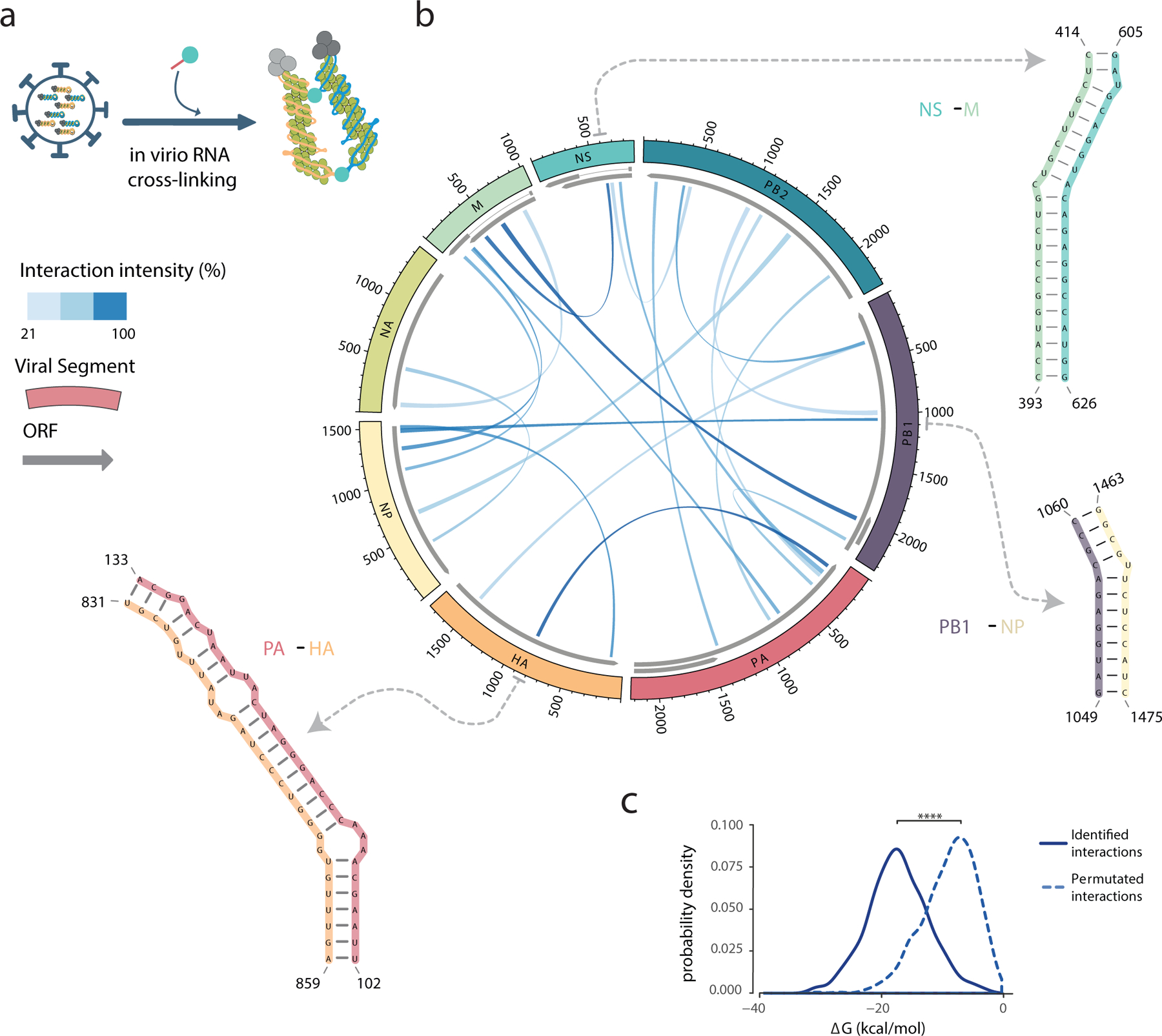
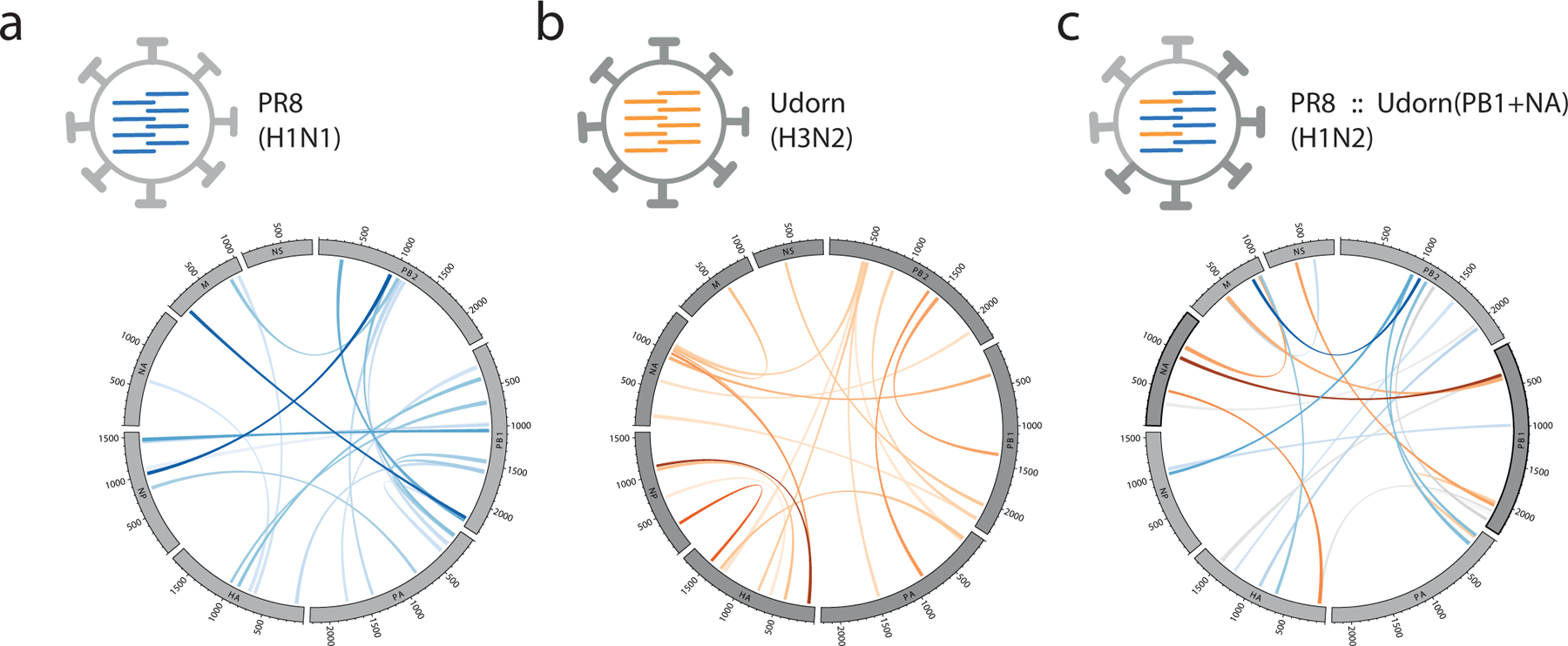
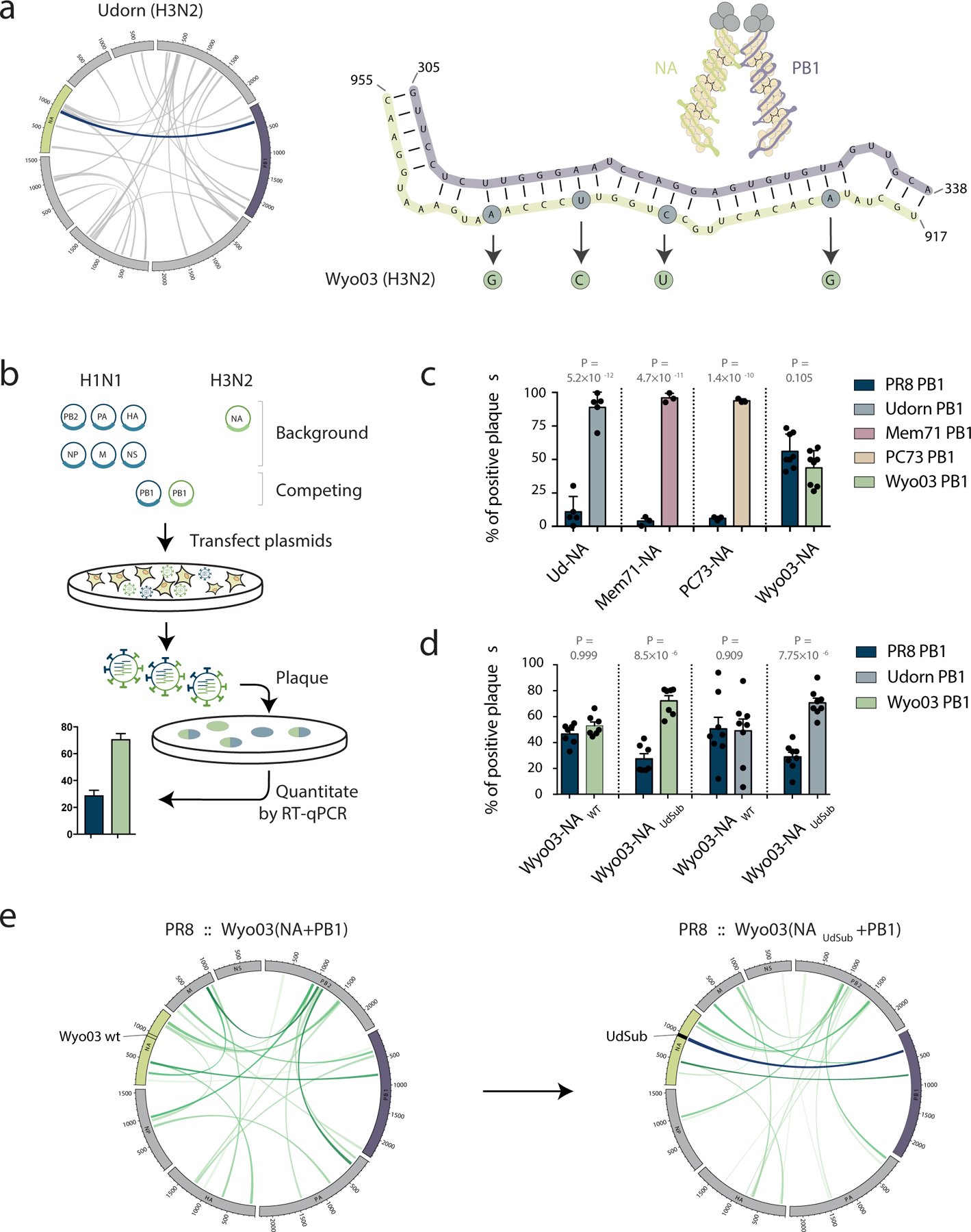
Influenza A viruses (IAVs) constitute a major threat to human health. The IAV genome consists of eight single-stranded viral RNA segments contained in separate viral ribonucleoprotein (vRNP) complexes that are packaged together into a single virus particle. The structure of viral RNA is believed to play a role in assembling the different vRNPs into budding virions1-8 and in directing reassortment between IAVs9. Reassortment between established human IAVs and IAVs harboured in the animal reservoir can lead to the emergence of pandemic influenza strains to which there is little pre-existing immunity in the human population10,11. While previous studies have revealed the overall organization of the proteins within vRNPs, characterization of viral RNA structure using conventional structural methods is hampered by limited resolution and an inability to resolve dynamic components12,13. Here, we employ multiple high-throughput sequencing approaches to generate a global high-resolution structure of the IAV genome. We show that different IAV genome segments acquire distinct RNA conformations and form both intra- and intersegment RNA interactions inside influenza virions. We use our detailed map of IAV genome structure to provide direct evidence for how intersegment RNA interactions drive vRNP cosegregation during reassortment between different IAV strains. The work presented here is a roadmap both for the development of improved vaccine strains and for the creation of a framework to 'risk assess' reassortment potential to better predict the emergence of new pandemic influenza strains.
Conflict of interest statement
Competing Interests
The authors declare no competing financial interests.
Figures
References
-
- Gerber M, Isel C, Moules V, Marquet R. Selective packaging of the influenza A genome and consequences for genetic reassortment. Trends Microbiol. 2014;22(8):446–455. - PubMed
-
- Hutchinson EC, von Kirchbach JC, Gog JR, Digard P. Genome packaging in influenza A virus. J Gen Virol. 2010;91(2):313–328. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
