

Geographic contrasts between pre- and postzygotic barriers are consistent with reinforcement in Heliconius butterflies
- PMID: 31334832
- PMCID: PMC6771877
- DOI: 10.1111/evo.13804
Geographic contrasts between pre- and postzygotic barriers are consistent with reinforcement in Heliconius butterflies
Abstract
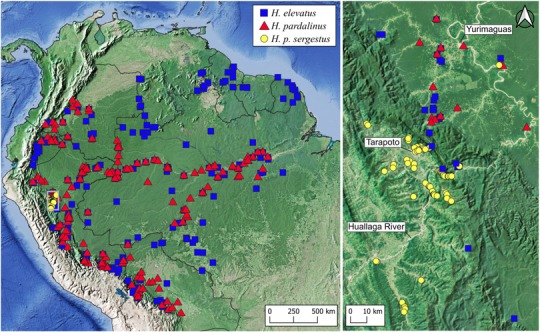
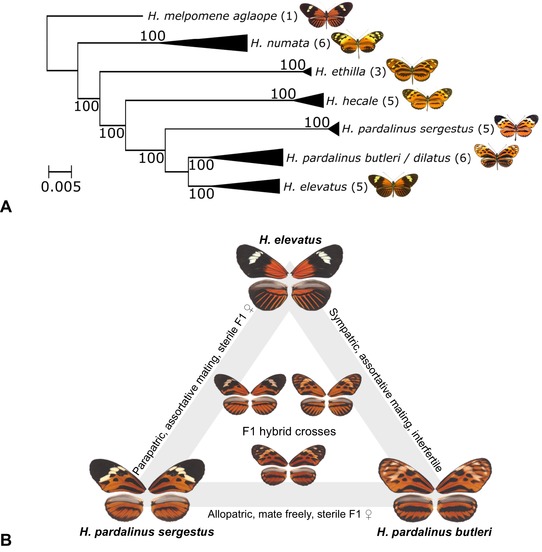
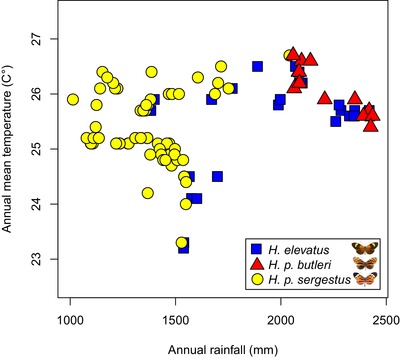
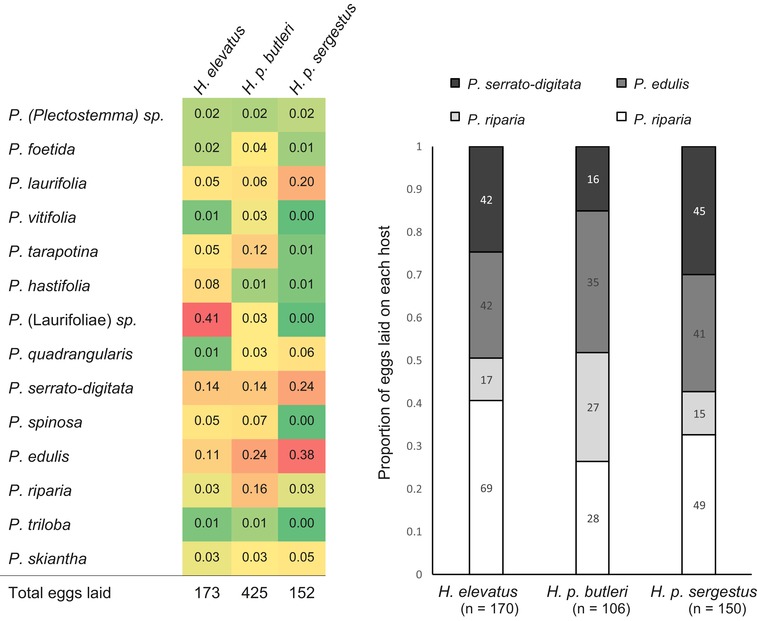
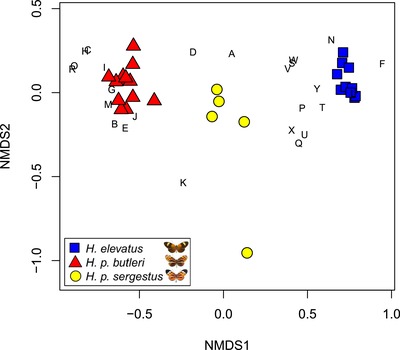
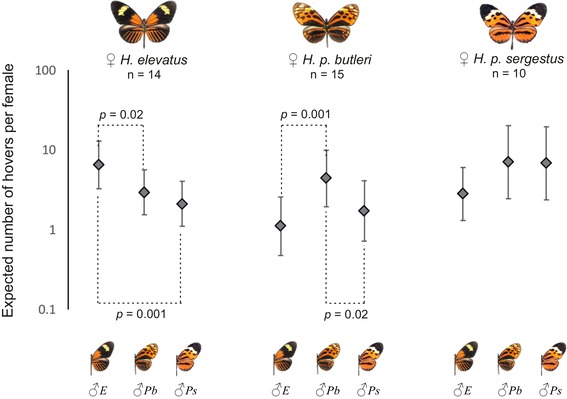
Identifying the traits causing reproductive isolation and the order in which they evolve is fundamental to understanding speciation. Here, we quantify prezygotic and intrinsic postzygotic isolation among allopatric, parapatric, and sympatric populations of the butterflies Heliconius elevatus and Heliconius pardalinus. Sympatric populations from the Amazon (H. elevatus and H. p. butleri) exhibit strong prezygotic isolation and rarely mate in captivity; however, hybrids are fertile. Allopatric populations from the Amazon (H. p. butleri) and Andes (H. p. sergestus) mate freely when brought together in captivity, but the female F1 hybrids are sterile. Parapatric populations (H. elevatus and H. p. sergestus) exhibit both assortative mating and sterility of female F1s. Assortative mating in sympatric populations is consistent with reinforcement in the face of gene flow, where the driving force, selection against hybrids, is due to disruption of mimicry and other ecological traits rather than hybrid sterility. In contrast, the lack of assortative mating and hybrid sterility observed in allopatric populations suggests that geographic isolation enables the evolution of intrinsic postzygotic reproductive isolation. Our results show how the types of reproductive barriers that evolve between species may depend on geography.
Keywords: Butterflies; gene flow; hybrid sterility; prezygotic isolation; speciation.
© 2019 The Authors. Evolution published by Wiley Periodicals, Inc. on behalf of The Society for the Study of Evolution.
Figures
References
-
- Arias, C. F. , Muñoz A. G., Jiggins C. D., Mavárez J., Bermingham E., and Linares M.. 2008. A hybrid zone provides evidence for incipient ecological speciation in Heliconius butterflies. Mol. Ecol. 17:4699–4712. - PubMed
-
- Barraclough, T. G. , and Vogler A. P.. 2000. Detecting the geographical pattern of speciation from species‐level phylogenies. Am. Nat. 155:419–434. - PubMed
-
- Bates, D. , Maechler M., Bolker B., and Walker S.. 2015. Fitting linear mixed‐effects models using lme4. J. Stat. Softw. 67:1–48.