

A-to-I editing of Malacoherpesviridae RNAs supports the antiviral role of ADAR1 in mollusks
- PMID: 31337330
- PMCID: PMC6651903
- DOI: 10.1186/s12862-019-1472-6
A-to-I editing of Malacoherpesviridae RNAs supports the antiviral role of ADAR1 in mollusks
Abstract
Background: Adenosine deaminase enzymes of the ADAR family are conserved in metazoans. They convert adenine into inosine in dsRNAs and thus alter both structural properties and the coding potential of their substrates. Acting on exogenous dsRNAs, ADAR1 exerts a pro- or anti-viral role in vertebrates and Drosophila.
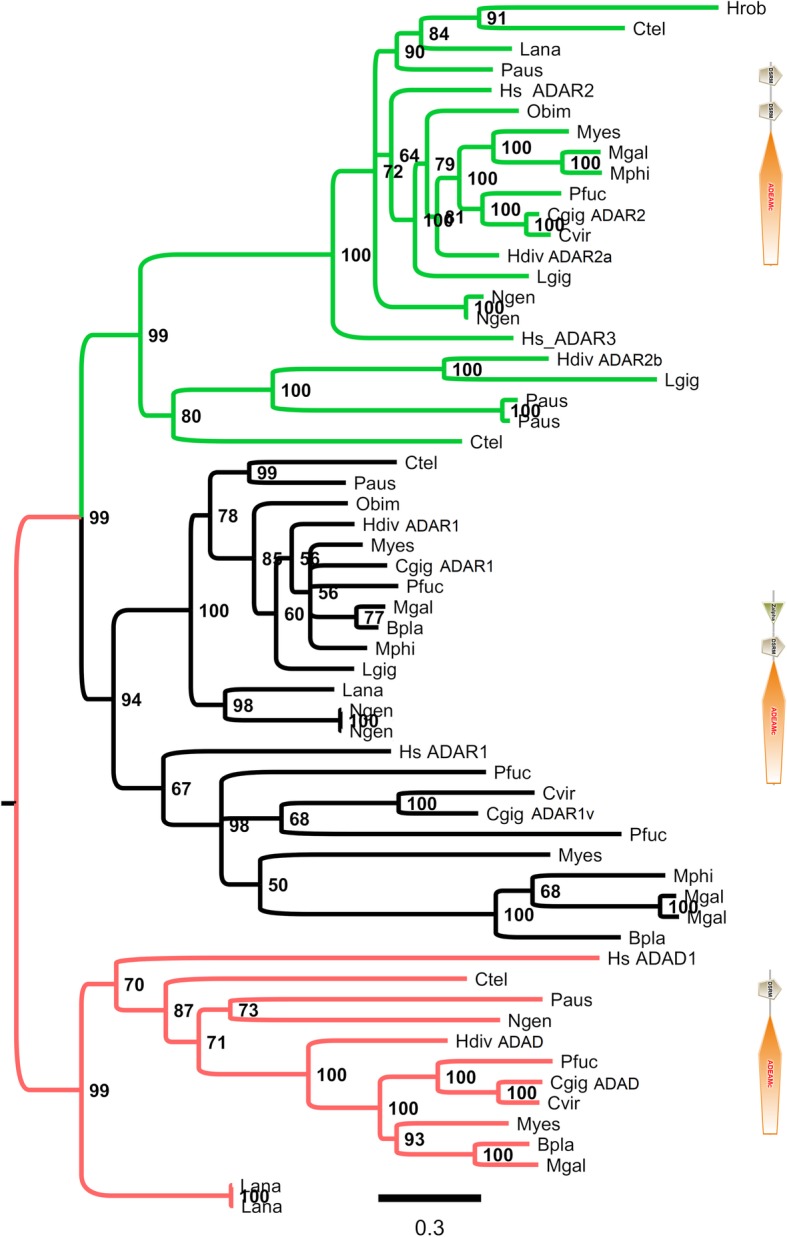
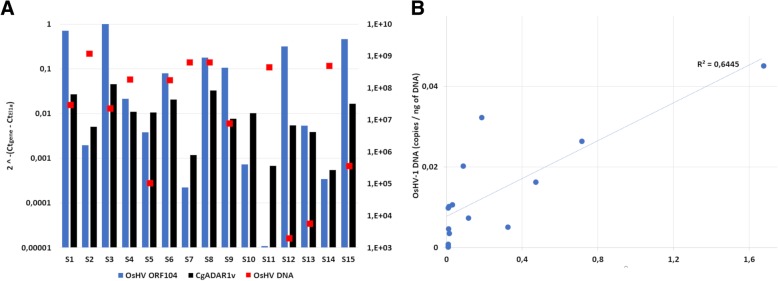
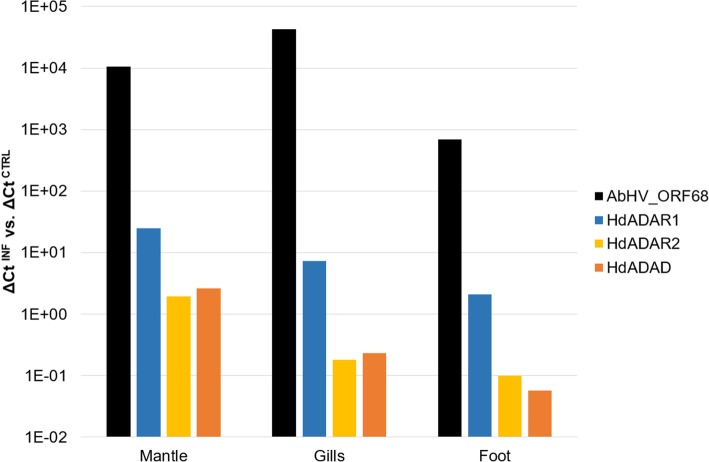
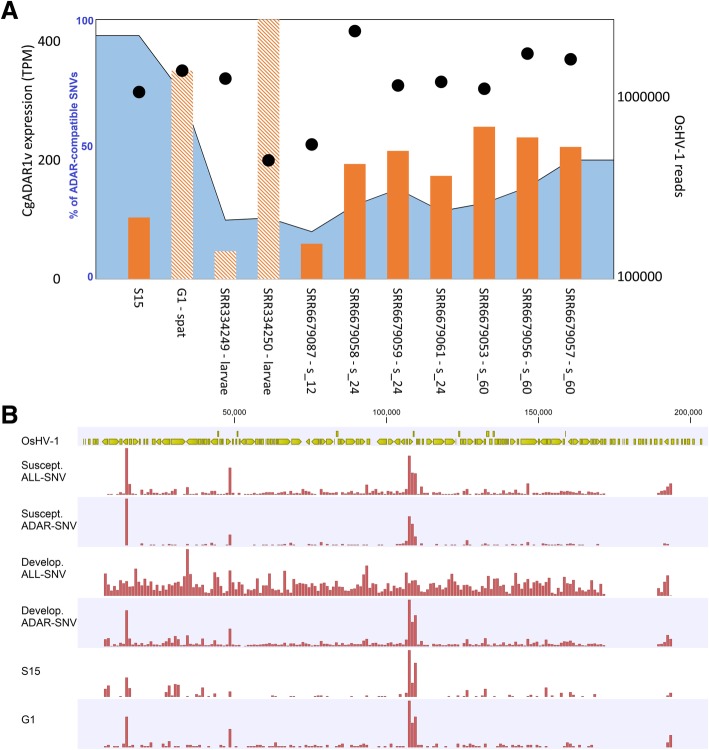
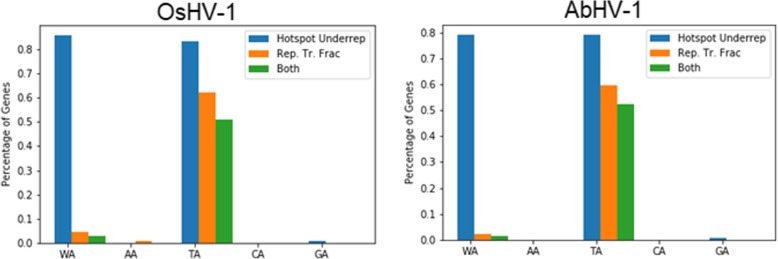
Results: We traced 4 ADAR homologs in 14 lophotrochozoan genomes and we classified them into ADAD, ADAR1 or ADAR2, based on phylogenetic and structural analyses of the enzymatic domain. Using RNA-seq and quantitative real time PCR we demonstrated the upregulation of one ADAR1 homolog in the bivalve Crassostrea gigas and in the gastropod Haliotis diversicolor supertexta during Ostreid herpesvirus-1 or Haliotid herpesvirus-1 infection. Accordingly, we demonstrated an extensive ADAR-mediated editing of viral RNAs. Single nucleotide variation (SNV) profiles obtained by pairing RNA- and DNA-seq data from the viral infected individuals resulted to be mostly compatible with ADAR-mediated A-to-I editing (up to 97%). SNVs occurred at low frequency in genomic hotspots, denoted by the overlapping of viral genes encoded on opposite DNA strands. The SNV sites and their upstream neighbor nucleotide indicated the targeting of selected adenosines. The analysis of viral sequences suggested that, under the pressure of the ADAR editing, the two Malacoherpesviridae genomes have evolved to reduce the number of deamination targets.
Conclusions: We report, for the first time, evidence of an extensive editing of Malacoherpesviridae RNAs attributable to host ADAR1 enzymes. The analysis of base neighbor preferences, structural features and expression profiles of molluscan ADAR1 supports the conservation of the enzyme function among metazoans and further suggested that ADAR1 exerts an antiviral role in mollusks.
Keywords: A-to-I editing; ADAR; AbHV-1; Abalones; Antiviral responses; Malacoherpesvirus; Mollusks; OsHV-1; Oysters; RNA editing.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
