

Genomic Evidence for Simultaneous Optimization of Transcription and Translation through Codon Variants in the pmoCAB Operon of Type Ia Methanotrophs
- PMID: 31337658
- PMCID: PMC6650546
- DOI: 10.1128/mSystems.00342-19
Genomic Evidence for Simultaneous Optimization of Transcription and Translation through Codon Variants in the pmoCAB Operon of Type Ia Methanotrophs
Abstract
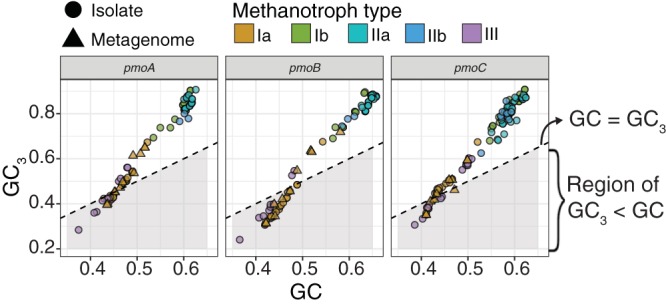
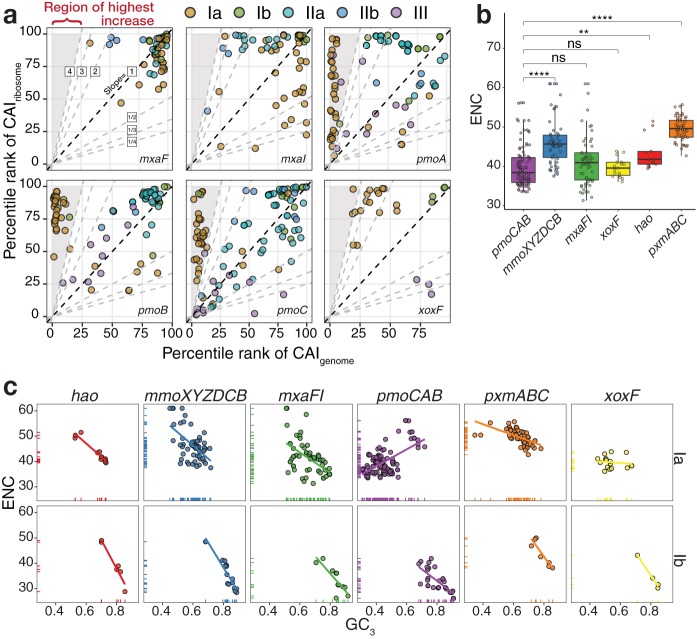
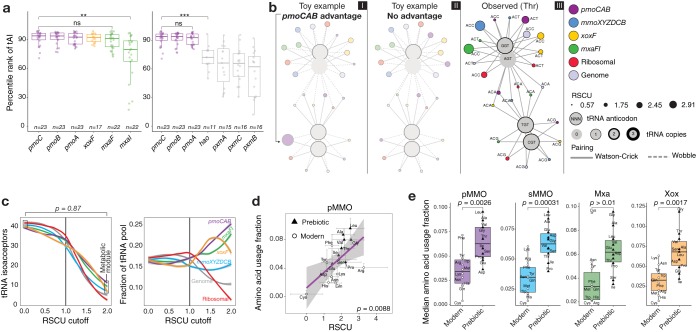
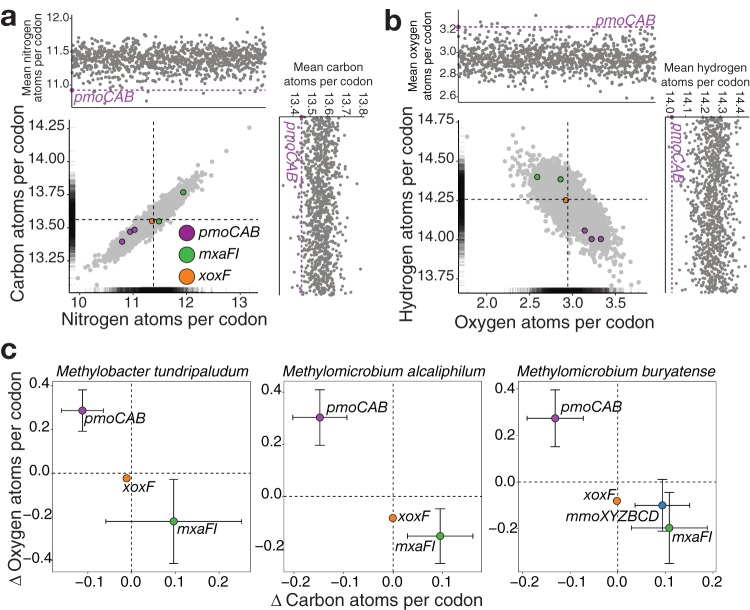
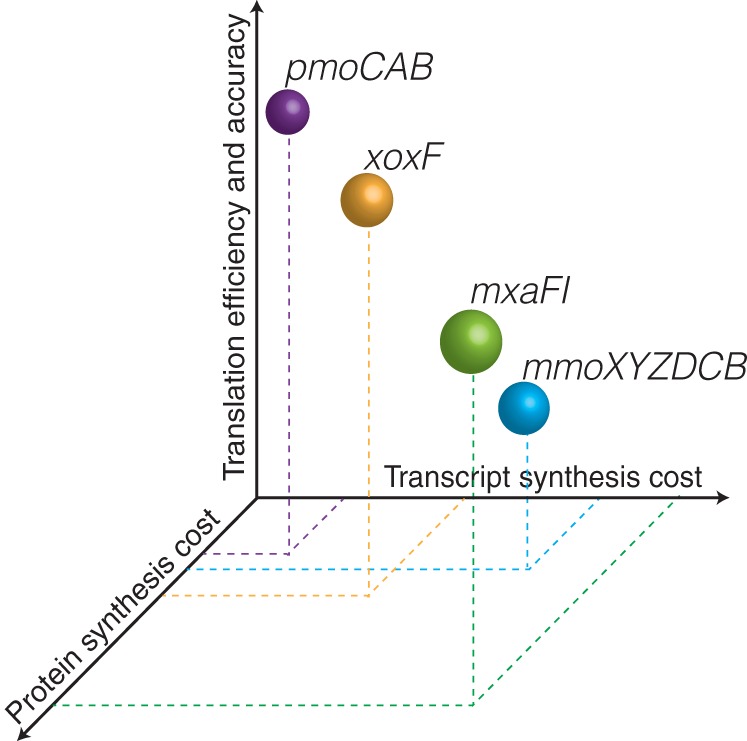
Understanding the interplay between genotype and phenotype is a fundamental goal of functional genomics. Methane oxidation is a microbial phenotype with global-scale significance as part of the carbon biogeochemical cycle and a sink for greenhouse gas. Microorganisms that oxidize methane (methanotrophs) are taxonomically diverse and widespread around the globe. In methanotrophic bacteria, enzymes in the methane oxidation metabolic module (KEGG module M00174, conversion of methane to formaldehyde) are encoded in four operons (pmoCAB, mmoXYZBCD, mxaFI, and xoxF). Recent reports have suggested that methanotrophs in Proteobacteria acquired methane monooxygenases through horizontal gene transfer. Here, we used a genomic meta-analysis to infer the transcriptional and translational advantages of coding sequences from the methane oxidation metabolic modules of different types of methanotrophs. By analyzing isolate and metagenome-assembled genomes from phylogenetically and geographically diverse sources, we detected an anomalous nucleotide composition bias in the coding sequences of particulate methane monooxygenase genes (pmoCAB) from type Ia methanotrophs. We found that this nucleotide bias increases the level of codon bias by decreasing the GC content in the third base of codons, a strategy that contrasts with that of other coding sequences in the module. Further codon usage analyses uncovered that codon variants of the type Ia pmoCAB coding sequences deviate from the genomic signature to match ribosomal protein-coding sequences. Subsequently, computation of transcription and translation metrics revealed that the pmoCAB coding sequences of type Ia methanotrophs optimize the usage of codon variants to maximize translation efficiency and accuracy, while minimizing the synthesis cost of transcripts and proteins.IMPORTANCE Microbial methane oxidation plays a fundamental role in the biogeochemical cycle of Earth's system. Recent reports have provided evidence for the acquisition of methane monooxygenases by horizontal gene transfer in methane-oxidizing bacteria from different environments, but how evolution has shaped the coding sequences to execute methanotrophy efficiently remains unexplored. In this work, we provide genomic evidence that among the different types of methanotrophs, type Ia methanotrophs possess a unique coding sequence of the pmoCAB operon that is under positive selection for optimal resource allocation and efficient synthesis of transcripts and proteins. This adaptive trait possibly enables type Ia methanotrophs to respond robustly to fluctuating methane availability and explains their global prevalence.
Keywords: codon usage; methane monooxygenase; resource allocation; synthesis cost; translation efficiency.
Copyright © 2019 Villada et al.
Figures
References
-
- Dean JF, Middelburg JJ, Röckmann T, Aerts R, Blauw LG, Egger M, Jetten MS, Jong AE, Meisel OH, Rasigraf O, Slomp CP, in't Zandt MH, Dolman AJ. 2018. Methane feedbacks to the global climate system in a warmer world. Rev Geophys 56:207–250. doi: 10.1002/2017RG000559. - DOI
-
- Oswald K, Milucka J, Brand A, Hach P, Littmann S, Wehrli B, Kuypers MM, Schubert CJ. 2016. Aerobic gammaproteobacterial methanotrophs mitigate methane emissions from oxic, and anoxic lake waters. Limnol Oceanogr 61:S101–S118. doi: 10.1002/lno.10312. - DOI
-
- Michaud AB, Dore JE, Achberger AM, Christner BC, Mitchell AC, Skidmore ML, Vick-Majors TJ, Priscu JC. 2017. Microbial oxidation as a methane sink beneath the West Antarctic Ice Sheet. Nature Geosci 10:582–586. doi: 10.1038/ngeo2992. - DOI
-
- Knoblauch C, Beer C, Liebner S, Grigoriev MN, Pfeiffer EM. 2018. Methane production as key to the greenhouse gas budget of thawing permafrost. Nat Clim Chang 8:309–312. doi: 10.1038/s41558-018-0095-z. - DOI
-
- Brankovits D, Pohlman JW, Niemann H, Leigh MB, Leewis MC, Becker KW, Iliffe TM, Alvarez F, Lehmann MF, Phillips B. 2017. Methane- and dissolved organic carbon-fueled microbial loop supports a tropical subterranean estuary ecosystem. Nat Commun 8:1835. doi: 10.1038/s41467-017-01776-x. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
