

Trading amino acids at the aphid- Buchnera symbiotic interface
- PMID: 31337682
- PMCID: PMC6690024
- DOI: 10.1073/pnas.1906223116
Trading amino acids at the aphid- Buchnera symbiotic interface
Abstract
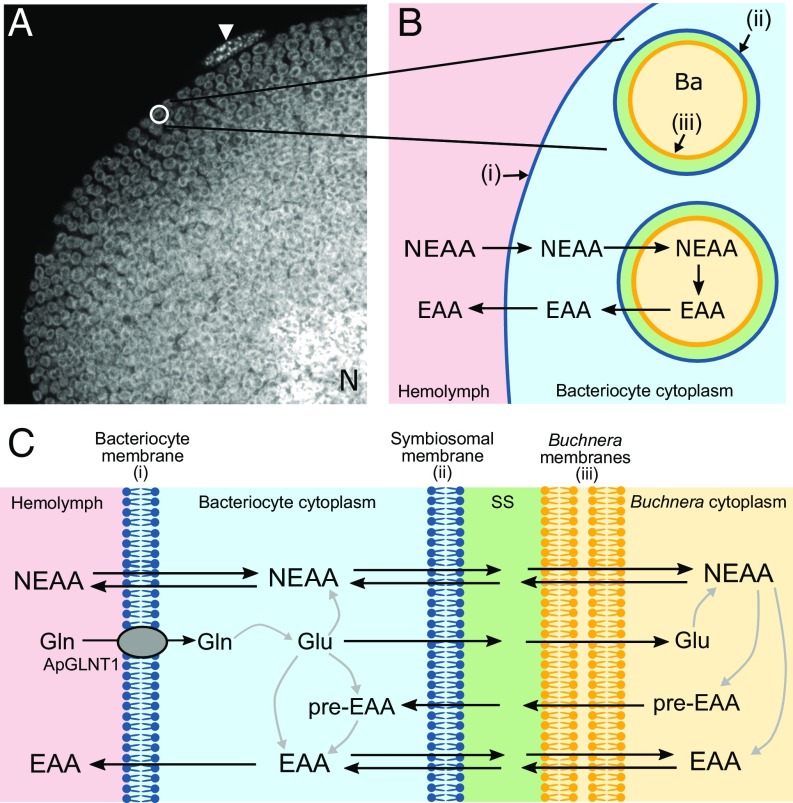
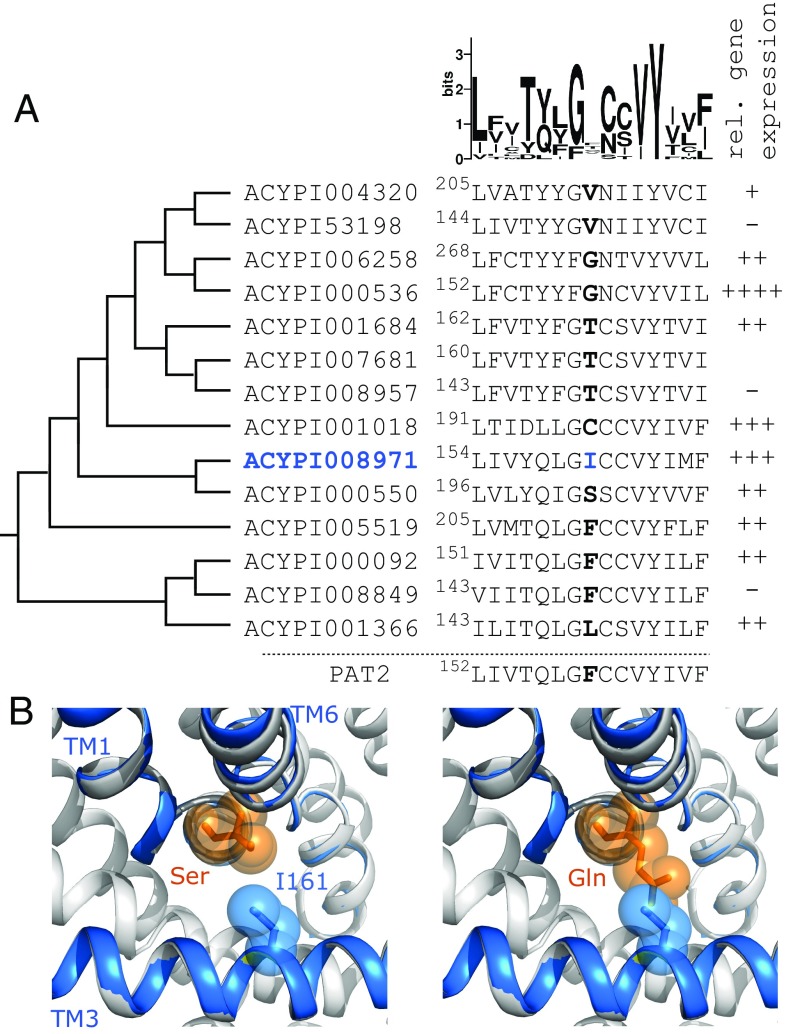
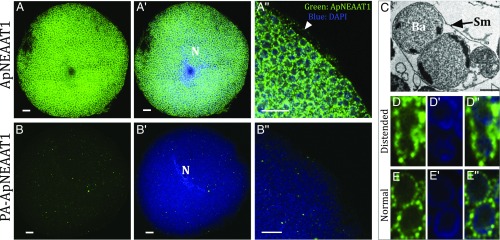
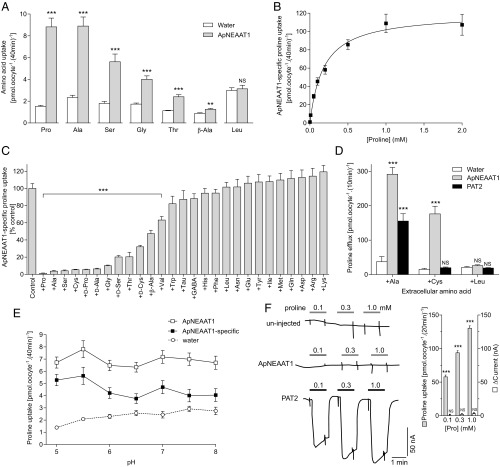
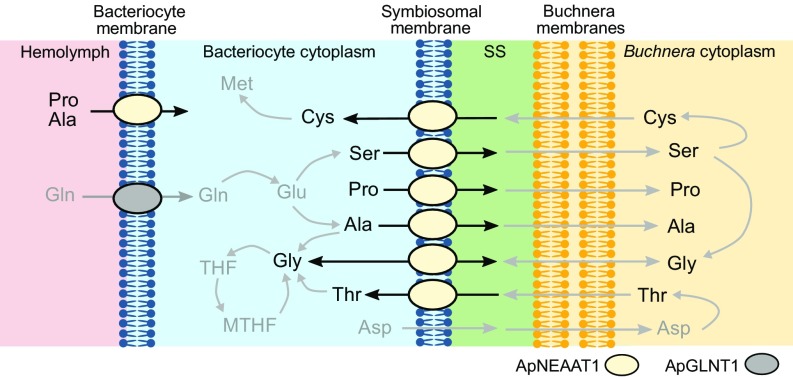
Plant sap-feeding insects are widespread, having evolved to occupy diverse environmental niches despite exclusive feeding on an impoverished diet lacking in essential amino acids and vitamins. Success depends exquisitely on their symbiotic relationships with microbial symbionts housed within specialized eukaryotic bacteriocyte cells. Each bacteriocyte is packed with symbionts that are individually surrounded by a host-derived symbiosomal membrane representing the absolute host-symbiont interface. The symbiosomal membrane must be a dynamic and selectively permeable structure to enable bidirectional and differential movement of essential nutrients, metabolites, and biosynthetic intermediates, vital for growth and survival of host and symbiont. However, despite this crucial role, the molecular basis of membrane transport across the symbiosomal membrane remains unresolved in all bacteriocyte-containing insects. A transport protein was immunolocalized to the symbiosomal membrane separating the pea aphid Acyrthosiphon pisum from its intracellular symbiont Buchnera aphidicola The transporter, A. pisum nonessential amino acid transporter 1, or ApNEAAT1 (gene: ACYPI008971), was characterized functionally following heterologous expression in Xenopus oocytes, and mediates both inward and outward transport of small dipolar amino acids (serine, proline, cysteine, alanine, glycine). Electroneutral ApNEAAT1 transport is driven by amino acid concentration gradients and is not coupled to transmembrane ion gradients. Previous metabolite profiling of hemolymph and bacteriocyte, alongside metabolic pathway analysis in host and symbiont, enable prediction of a physiological role for ApNEAAT1 in bidirectional host-symbiont amino acid transfer, supplying both host and symbiont with indispensable nutrients and biosynthetic precursors to facilitate metabolic complementarity.
Keywords: amino acid transport; metabolic integration; symbiosis.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Douglas A. E., The Symbiotic Habit (Princeton University Press, 2010).
-
- DeSalle R., Perkins S. L., Welcome to the Microbiome: Getting to Know the Trillions of Bacteria and Other Microbes in, on, and Around You (Yale University Press, 2015).
-
- Yong E., I Contain Multitudes: The Microbes within Us and a Grander View of Life (HarperCollins, 2016).
-
- Moran N. A., McCutcheon J. P., Nakabachi A., Genomics and evolution of heritable bacterial symbionts. Annu. Rev. Genet. 42, 165–190 (2008). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
