

Wheat VRN1, FUL2 and FUL3 play critical and redundant roles in spikelet development and spike determinacy
- PMID: 31337701
- PMCID: PMC6679363
- DOI: 10.1242/dev.175398
Wheat VRN1, FUL2 and FUL3 play critical and redundant roles in spikelet development and spike determinacy
Abstract
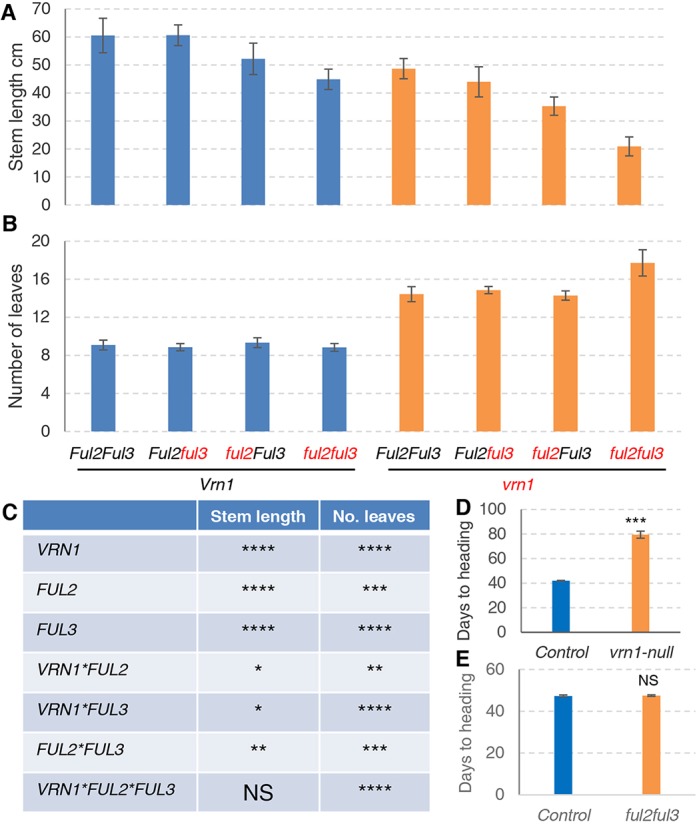
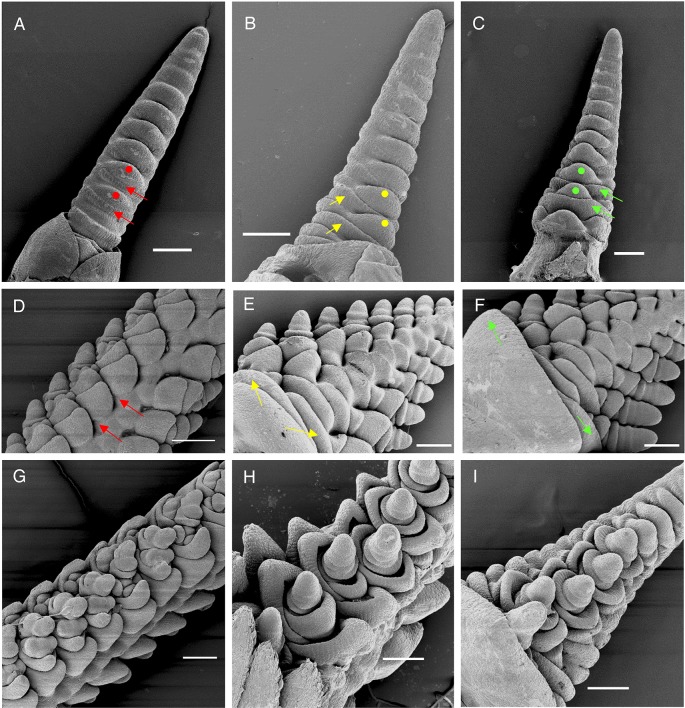
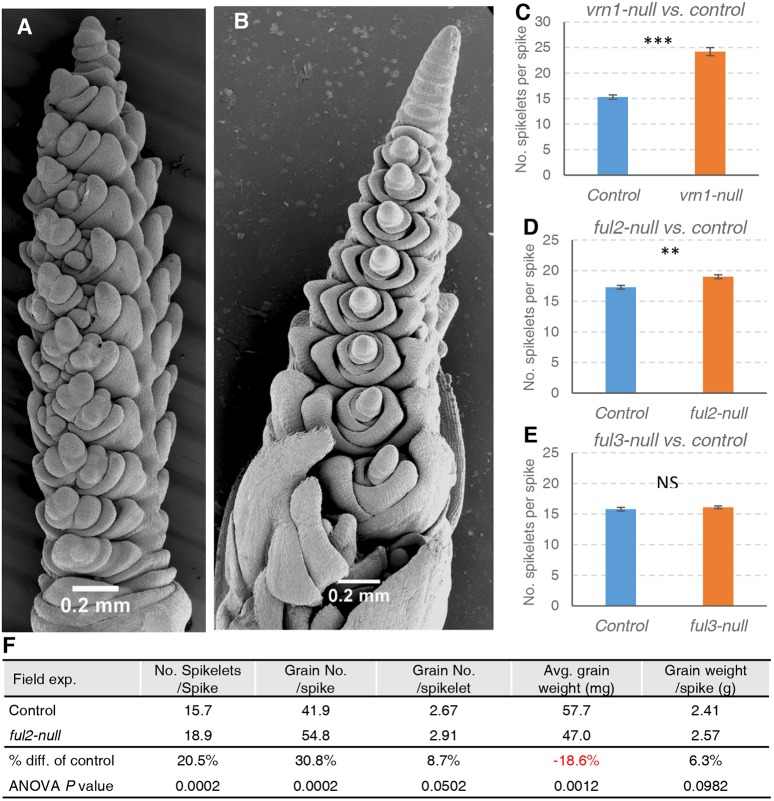
The spikelet is the basic unit of the grass inflorescence. In this study, we show that wheat MADS-box genes VRN1, FUL2 and FUL3 play critical and redundant roles in spikelet and spike development, and also affect flowering time and plant height. In the vrn1ful2ful3-null triple mutant, the inflorescence meristem formed a normal double-ridge structure, but then the lateral meristems generated vegetative tillers subtended by leaves instead of spikelets. These results suggest an essential role of these three genes in the fate of the upper spikelet ridge and the suppression of the lower leaf ridge. Inflorescence meristems of vrn1ful2ful3-null and vrn1ful2-null remained indeterminate and single vrn1-null and ful2-null mutants showed delayed formation of the terminal spikelet and increased number of spikelets per spike. Moreover, the ful2-null mutant showed more florets per spikelet, which together with a higher number of spikelets, resulted in a significant increase in the number of grains per spike in the field. Our results suggest that a better understanding of the mechanisms underlying wheat spikelet and spike development can inform future strategies to improve grain yield in wheat.
Keywords: MADS-box; Meristem identity; Spike development; Spikelet; Wheat.
© 2019. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures

Similar articles
-
APETALA 2-like genes AP2L2 and Q specify lemma identity and axillary floral meristem development in wheat.Plant J. 2020 Jan;101(1):171-187. doi: 10.1111/tpj.14528. Epub 2019 Oct 15. Plant J. 2020. PMID: 31494998 Free PMC article.
-
LEAFY and WAPO1 jointly regulate spikelet number per spike and floret development in wheat.Development. 2024 Aug 1;151(15):dev202803. doi: 10.1242/dev.202803. Epub 2024 Jul 31. Development. 2024. PMID: 39082949 Free PMC article.
-
Interactions between SQUAMOSA and SHORT VEGETATIVE PHASE MADS-box proteins regulate meristem transitions during wheat spike development.Plant Cell. 2021 Dec 3;33(12):3621-3644. doi: 10.1093/plcell/koab243. Plant Cell. 2021. PMID: 34726755 Free PMC article.
-
Grass meristems II: inflorescence architecture, flower development and meristem fate.Plant Cell Physiol. 2013 Mar;54(3):313-24. doi: 10.1093/pcp/pct016. Epub 2013 Jan 31. Plant Cell Physiol. 2013. PMID: 23378448 Review.
-
Molecular Insights into Inflorescence Meristem Specification for Yield Potential in Cereal Crops.Int J Mol Sci. 2021 Mar 29;22(7):3508. doi: 10.3390/ijms22073508. Int J Mol Sci. 2021. PMID: 33805287 Free PMC article. Review.
Cited by
-
APETALA 2-like genes AP2L2 and Q specify lemma identity and axillary floral meristem development in wheat.Plant J. 2020 Jan;101(1):171-187. doi: 10.1111/tpj.14528. Epub 2019 Oct 15. Plant J. 2020. PMID: 31494998 Free PMC article.
-
Identification and characterization of a natural polymorphism in FT-A2 associated with increased number of grains per spike in wheat.Theor Appl Genet. 2022 Feb;135(2):679-692. doi: 10.1007/s00122-021-03992-y. Epub 2021 Nov 26. Theor Appl Genet. 2022. PMID: 34825926 Free PMC article.
-
The Streptochaeta Genome and the Evolution of the Grasses.Front Plant Sci. 2021 Oct 4;12:710383. doi: 10.3389/fpls.2021.710383. eCollection 2021. Front Plant Sci. 2021. PMID: 34671369 Free PMC article.
-
INDETERMINATE1-mediated expression of FT family genes is required for proper timing of flowering in Brachypodium distachyon.Proc Natl Acad Sci U S A. 2023 Nov 14;120(46):e2312052120. doi: 10.1073/pnas.2312052120. Epub 2023 Nov 7. Proc Natl Acad Sci U S A. 2023. PMID: 37934817 Free PMC article.
-
TaAPO-A1, an ortholog of rice ABERRANT PANICLE ORGANIZATION 1, is associated with total spikelet number per spike in elite European hexaploid winter wheat (Triticum aestivum L.) varieties.Sci Rep. 2019 Sep 25;9(1):13853. doi: 10.1038/s41598-019-50331-9. Sci Rep. 2019. PMID: 31554871 Free PMC article.
