

LZTR1 facilitates polyubiquitination and degradation of RAS-GTPases
- PMID: 31337872
- PMCID: PMC7206011
- DOI: 10.1038/s41418-019-0395-5
LZTR1 facilitates polyubiquitination and degradation of RAS-GTPases
Abstract
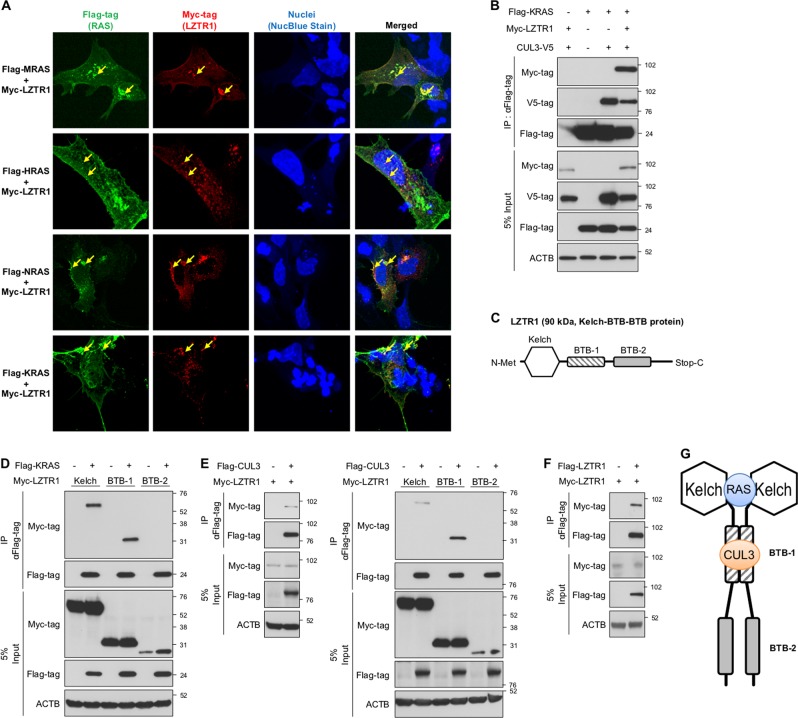
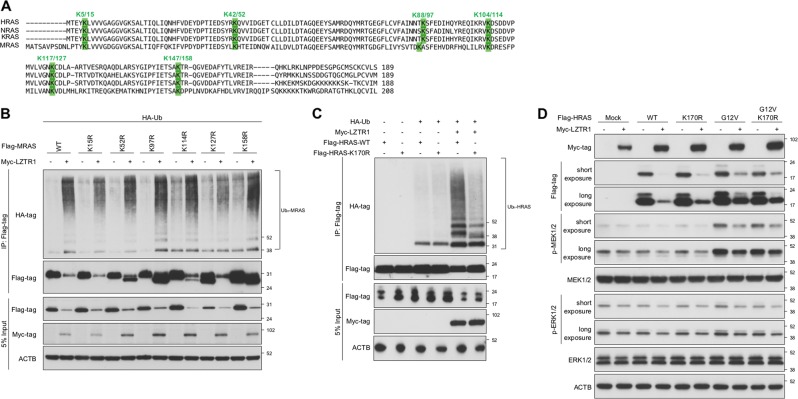
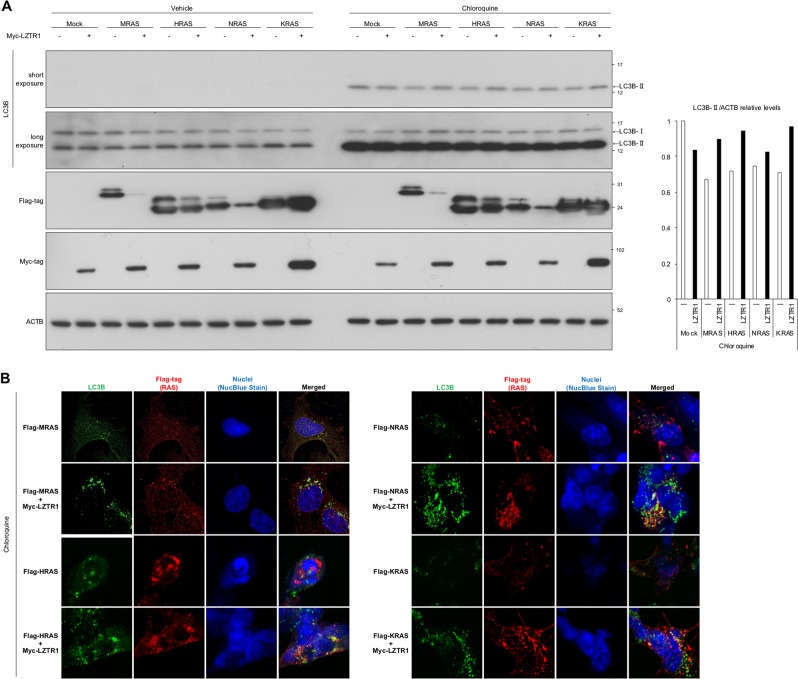
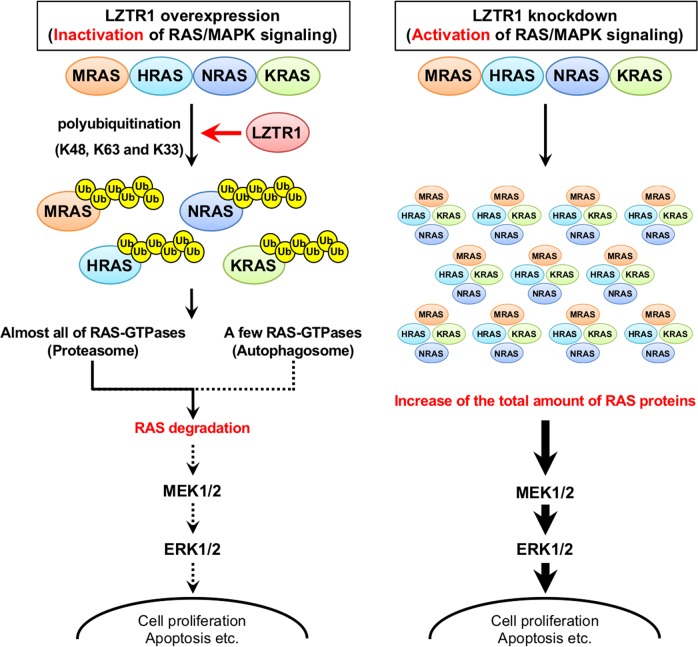
Leucine zipper-like transcriptional regulator 1 (LZTR1) encodes a member of the BTB-Kelch superfamily, which interacts with the Cullin3 (CUL3)-based E3 ubiquitin ligase complex. Mutations in LZTR1 have been identified in glioblastoma, schwannomatosis, and Noonan syndrome. However, the functional role of LZTR1 in carcinogenesis or human development is not fully understood. Here, we demonstrate that LZTR1 facilitates the polyubiquitination and degradation of RAS via the ubiquitin-proteasome pathway, leading to the inhibition of the RAS/MAPK signaling. The polyubiquitination and degradation of RAS was also observed in cells expressing MRAS, HRAS, NRAS, and KRAS as well as oncogenic RAS mutants and inhibited the activation of ERK1/2 and cell growth. In vivo ubiquitination assays showed that MRAS-K127 and HRAS-K170 were ubiquitinated by LZTR1 and that the polyubiquitinated-chains contained mainly Ub-K48, K63, and K33-linked chains, suggesting its possible involvement in autophagy. Immunoprecipitation analyses showed the interaction of LZTR1 and RAS-GTPases with autophagy-related proteins, including LC3B and SQSTM1/p62. Co-expression of LZTR1 and RAS increased the expression of lipidated form of LC3B. However, long-term treatment with chloroquine had little effect on RAS protein levels, suggesting that the contribution of autophagy to LZTR1-mediated RAS degradation is minimal. Taken together, these results show that LZTR1 functions as a "RAS killer protein" mainly via the ubiquitin-proteasome pathway regardless of the type of RAS GTPase, controlling downstream signal transduction. Our results also suggest a possible association of LZTR1 and RAS-GTPases with the autophagy. These findings provide clues for the elucidation of the mechanisms of RAS degradation and regulation of the RAS/MAPK signaling cascade.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Nacak TG, Leptien K, Fellner D, Augustin HG, Kroll J. The BTB-kelch protein LZTR-1 is a novel Golgi protein that is degraded upon induction of apoptosis. J Biol Chem. 2006;281:5065–71. - PubMed
-
- Furukawa M, He YJ, Borchers C, Xiong Y. Targeting of protein ubiquitination by BTB-Cullin 3-Roc1 ubiquitin ligases. Nat Cell Biol. 2003;5:1001–7. - PubMed
-
- Geyer R, Wee S, Anderson S, Yates J, Wolf DA. BTB/POZ domain proteins are putative substrate adaptors for cullin 3 ubiquitin ligases. Mol Cell. 2003;12:783–90. - PubMed
-
- Stogios PJ, Prive GG. The BACK domain in BTB-kelch proteins. Trends Biochem Sci. 2004;29:634–7. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
