

The 1918 influenza pandemic: 100 years of questions answered and unanswered
- PMID: 31341062
- PMCID: PMC11000447
- DOI: 10.1126/scitranslmed.aau5485
The 1918 influenza pandemic: 100 years of questions answered and unanswered
Abstract
The 2018-2019 period marks the centennial of the "Spanish" influenza pandemic, which caused at least 50 million deaths worldwide. The unprecedented nature of the pandemic's sudden appearance and high fatality rate serve as a stark reminder of the threat influenza poses. Unusual features of the 1918-1919 pandemic, including age-specific mortality and the high frequency of severe pneumonias, are still not fully understood. Sequencing and reconstruction of the 1918 virus has allowed scientists to answer many questions about its origin and pathogenicity, although many questions remain. This Review summarizes key findings and still-to-be answered questions about this deadliest of human events.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
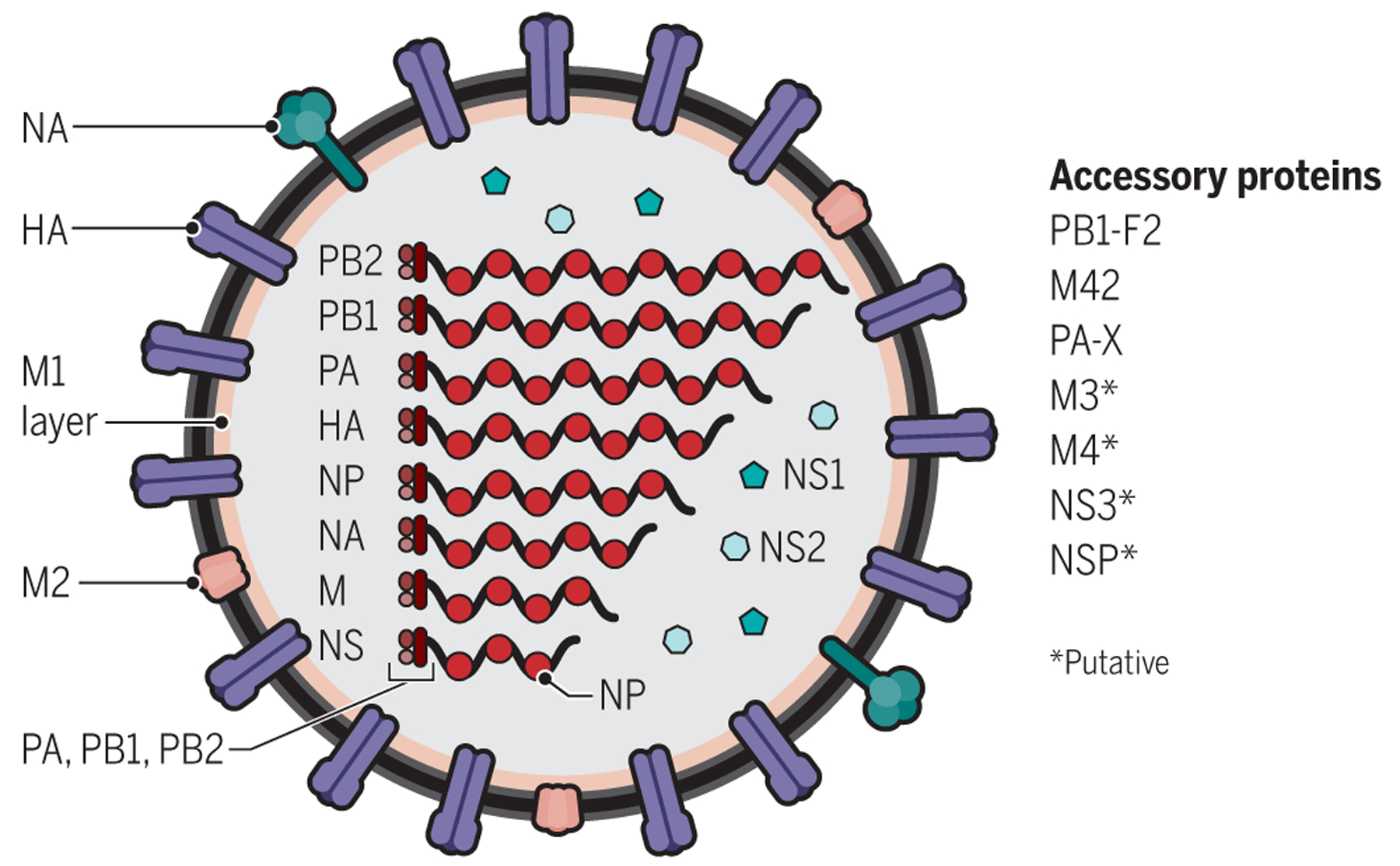




References
-
- Jordan EO, Epidemic Influenza: A Survey (American Medical Association, 1927).
-
- Chandra S, Kuljanin G, Wray J, Mortality from the influenza pandemic of 1918–1919: The case of India. Demography 49, 857–865 (2012). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
