

Lipidomics unveils lipid dyshomeostasis and low circulating plasmalogens as biomarkers in a monogenic mitochondrial disorder
- PMID: 31341105
- PMCID: PMC6675547
- DOI: 10.1172/jci.insight.123231
Lipidomics unveils lipid dyshomeostasis and low circulating plasmalogens as biomarkers in a monogenic mitochondrial disorder
Abstract
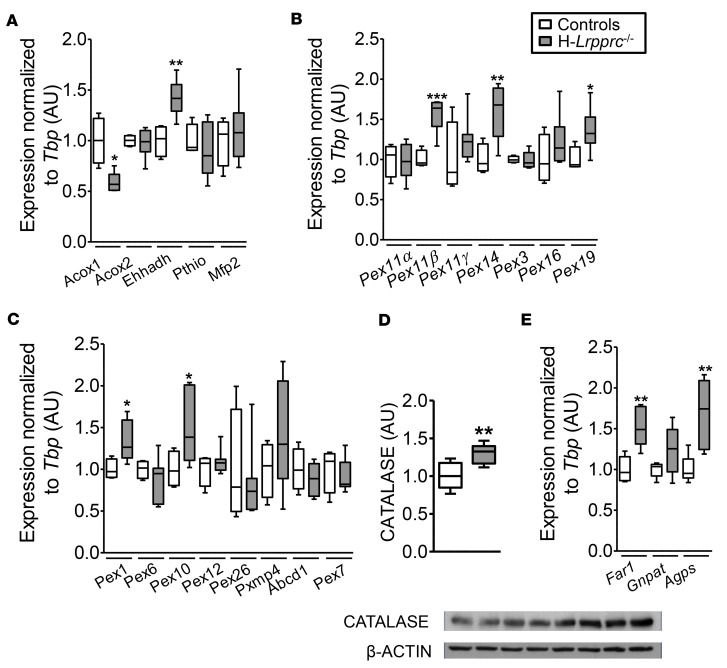
Mitochondrial dysfunction characterizes many rare and common age-associated diseases. The biochemical consequences, underlying clinical manifestations, and potential therapeutic targets, remain to be better understood. We tested the hypothesis that lipid dyshomeostasis in mitochondrial disorders goes beyond mitochondrial fatty acid β-oxidation, particularly in liver. This was achieved using comprehensive untargeted and targeted lipidomics in a case-control cohort of patients with Leigh syndrome French-Canadian variant (LSFC), a mitochondrial disease caused by mutations in LRPPRC, and in mice harboring liver-specific inactivation of Lrpprc (H-Lrpprc-/-). We discovered a plasma lipid signature discriminating LSFC patients from controls encompassing lower levels of plasmalogens and conjugated bile acids, which suggest perturbations in peroxisomal lipid metabolism. This premise was reinforced in H-Lrpprc-/- mice, which compared with littermates recapitulated a similar, albeit stronger peroxisomal metabolic signature in plasma and liver including elevated levels of very-long-chain acylcarnitines. These mice also presented higher transcript levels for hepatic markers of peroxisome proliferation in addition to lipid remodeling reminiscent of nonalcoholic fatty liver diseases. Our study underscores the value of lipidomics to unveil unexpected mechanisms underlying lipid dyshomeostasis ensuing from mitochondrial dysfunction herein implying peroxisomes and liver, which likely contribute to the pathophysiology of LSFC, but also other rare and common mitochondrial diseases.
Keywords: Cell Biology; Fatty acid oxidation; Metabolism; Mitochondria; Monogenic diseases.
Conflict of interest statement
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
