

Small RNA discovery in the interaction between barley and the powdery mildew pathogen
- PMID: 31345162
- PMCID: PMC6657096
- DOI: 10.1186/s12864-019-5947-z
Small RNA discovery in the interaction between barley and the powdery mildew pathogen
Erratum in
-
Correction to: small RNA discovery in the interaction between barley and the powdery mildew pathogen.BMC Genomics. 2019 Sep 4;20(1):697. doi: 10.1186/s12864-019-6012-7. BMC Genomics. 2019. PMID: 31484492 Free PMC article.
Abstract
Background: Plants encounter pathogenic and non-pathogenic microorganisms on a nearly constant basis. Small RNAs such as siRNAs and miRNAs/milRNAs influence pathogen virulence and host defense responses. We exploited the biotrophic interaction between the powdery mildew fungus, Blumeria graminis f. sp. hordei (Bgh), and its diploid host plant, barley (Hordeum vulgare) to explore fungal and plant sRNAs expressed during Bgh infection of barley leaf epidermal cells.
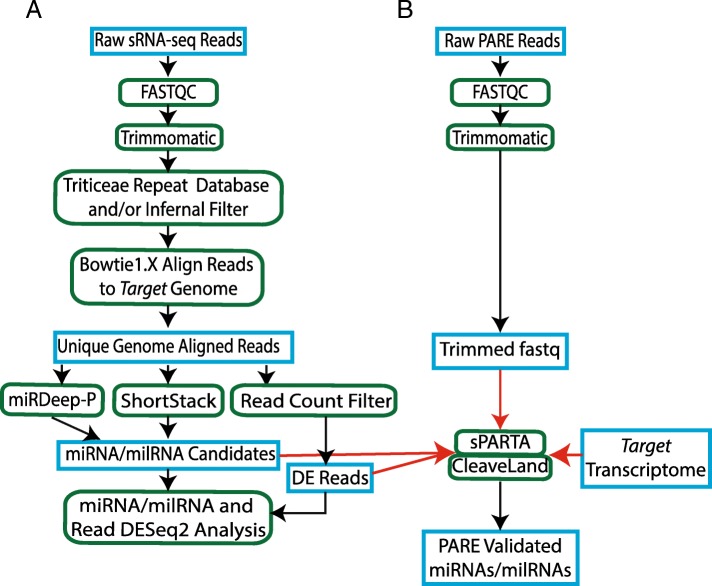
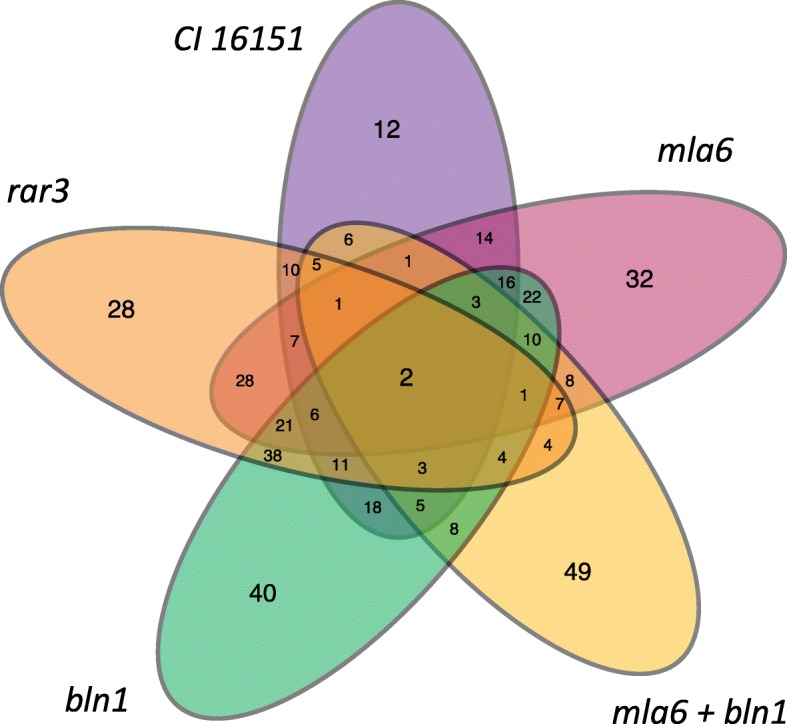
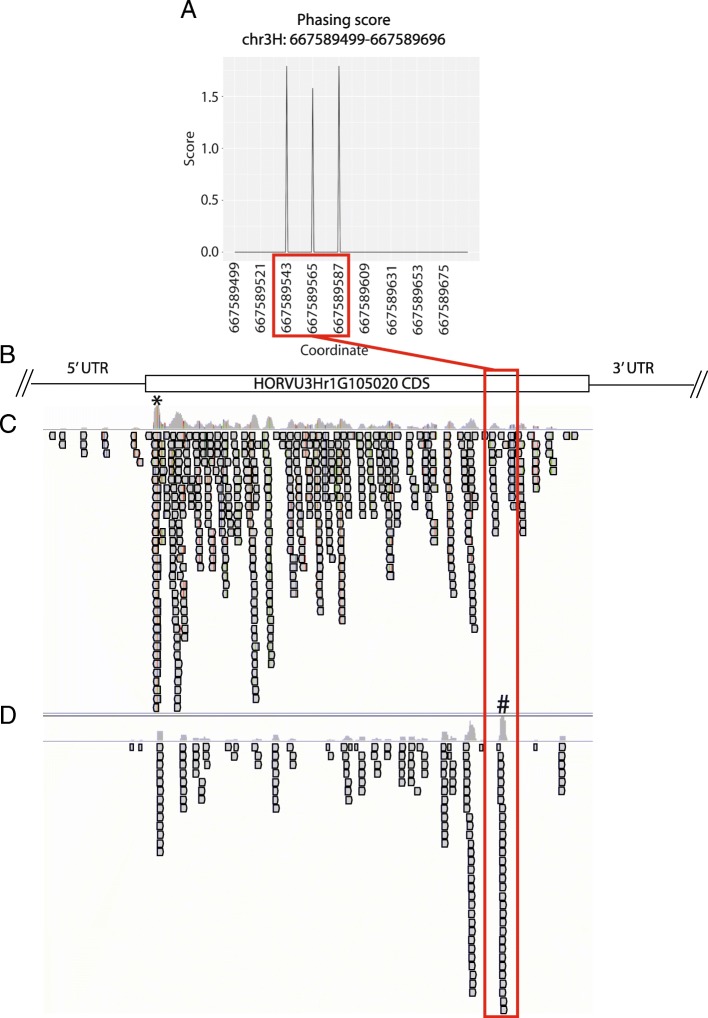
Results: RNA was isolated from four fast-neutron immune-signaling mutants and their progenitor over a time course representing key stages of Bgh infection, including appressorium formation, penetration of epidermal cells, and development of haustorial feeding structures. The Cereal Introduction (CI) 16151 progenitor carries the resistance allele Mla6, while Bgh isolate 5874 harbors the AVRa6 avirulence effector, resulting in an incompatible interaction. Parallel Analysis of RNA Ends (PARE) was used to verify sRNAs with likely transcript targets in both barley and Bgh. Bgh sRNAs are predicted to regulate effectors, metabolic genes, and translation-related genes. Barley sRNAs are predicted to influence the accumulation of transcripts that encode auxin response factors, NAC transcription factors, homeodomain transcription factors, and several splicing factors. We also identified phasing small interfering RNAs (phasiRNAs) in barley that overlap transcripts that encode receptor-like kinases (RLKs) and nucleotide-binding, leucine-rich domain proteins (NLRs).
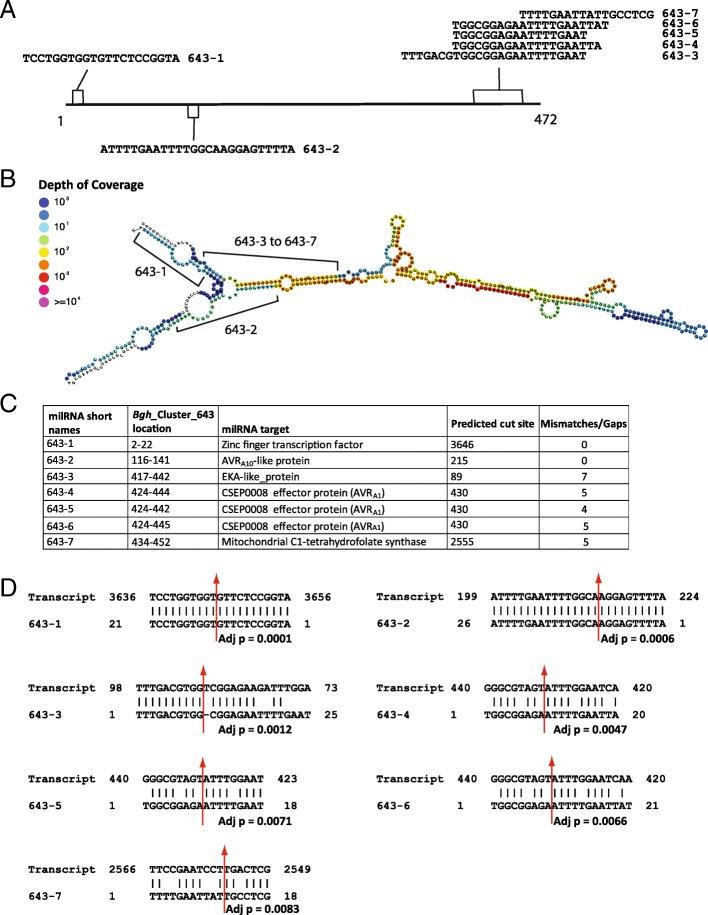
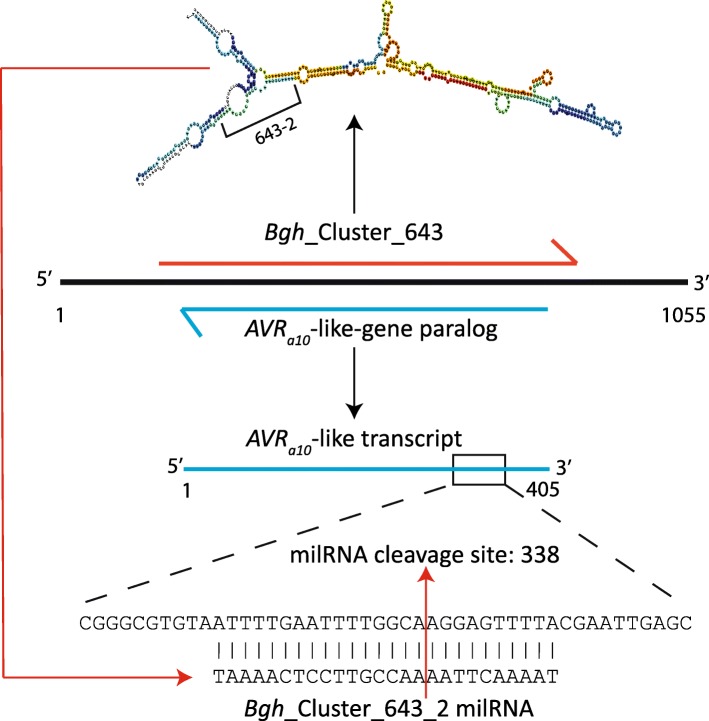
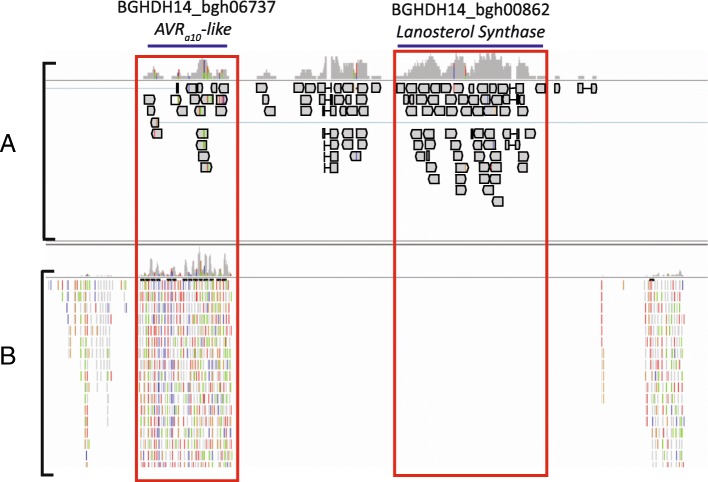
Conclusions: These data suggest that Bgh sRNAs regulate gene expression in metabolism, translation-related, and pathogen effectors. PARE-validated targets of predicted Bgh milRNAs include both EKA (effectors homologous to AVRk1 and AVRa10) and CSEP (candidate secreted effector protein) families. We also identified barley phasiRNAs and miRNAs in response to Bgh infection. These include phasiRNA loci that overlap with a significant proportion of receptor-like kinases, suggesting an additional sRNA control mechanism may be active in barley leaves as opposed to predominant R-gene phasiRNA overlap in many eudicots. In addition, we identified conserved miRNAs, novel miRNA candidates, and barley genome mapped sRNAs that have PARE validated transcript targets in barley. The miRNA target transcripts are enriched in transcription factors, signaling-related proteins, and photosynthesis-related proteins. Together these results suggest both barley and Bgh control metabolism and infection-related responses via the specific accumulation and targeting of genes via sRNAs.
Keywords: Barley; Blumeria; CSEPs; EKA family; Pathogen effectors; Small RNA-Seq; Transposable elements.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Amselem J, Vigouroux M, Oberhaensli S, Brown JK, Bindschedler LV, Skamnioti P, Wicker T, Spanu PD, Quesneville H, Sacristan S. Evolution of the EKA family of powdery mildew avirulence-effector genes from the ORF 1 of a LINE retrotransposon. BMC Genomics. 2015;16:917. doi: 10.1186/s12864-015-2185-x. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
