

Detection and activation of HIV broadly neutralizing antibody precursor B cells using anti-idiotypes
- PMID: 31345930
- PMCID: PMC6780997
- DOI: 10.1084/jem.20190164
Detection and activation of HIV broadly neutralizing antibody precursor B cells using anti-idiotypes
Abstract
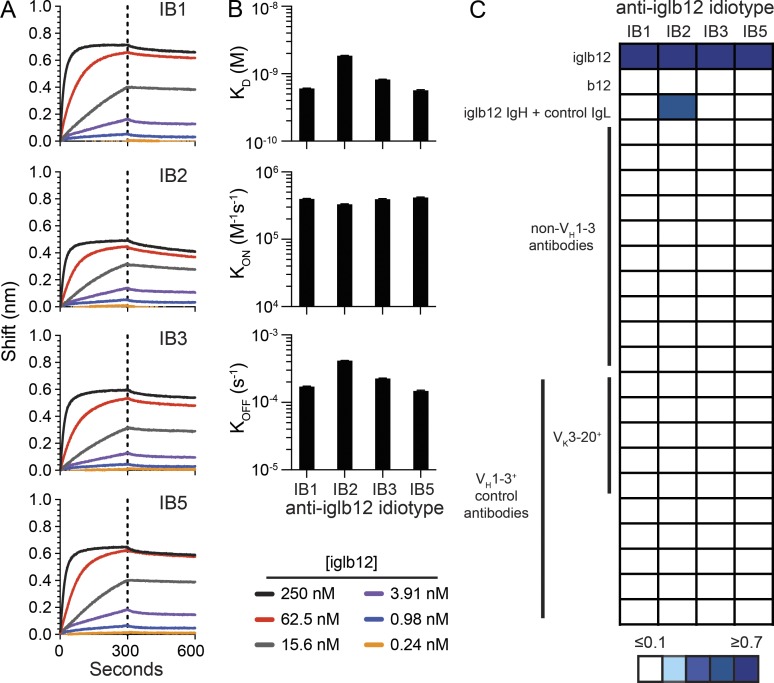
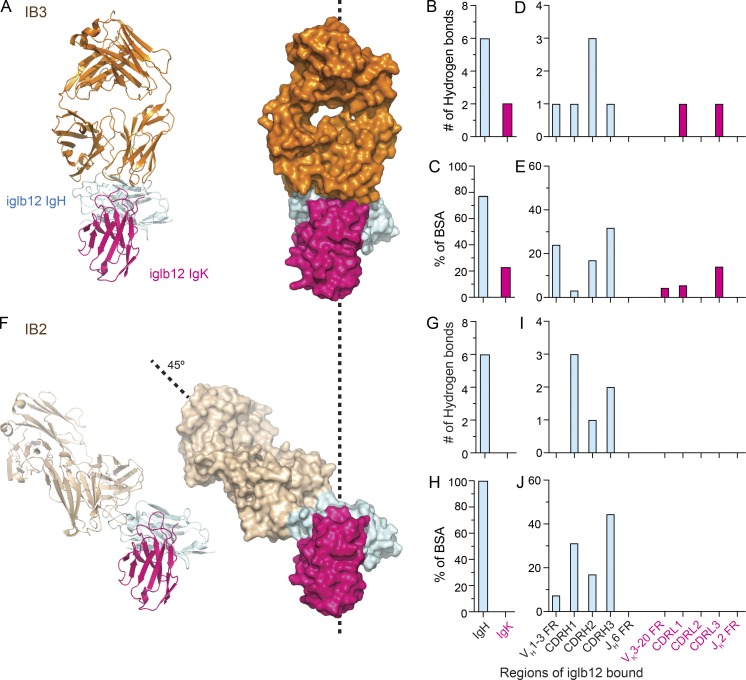
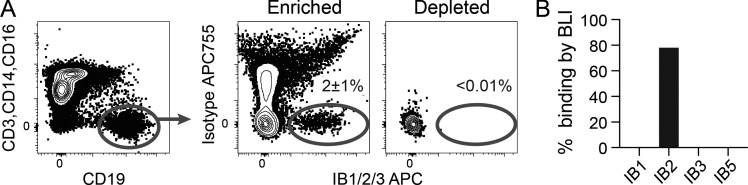
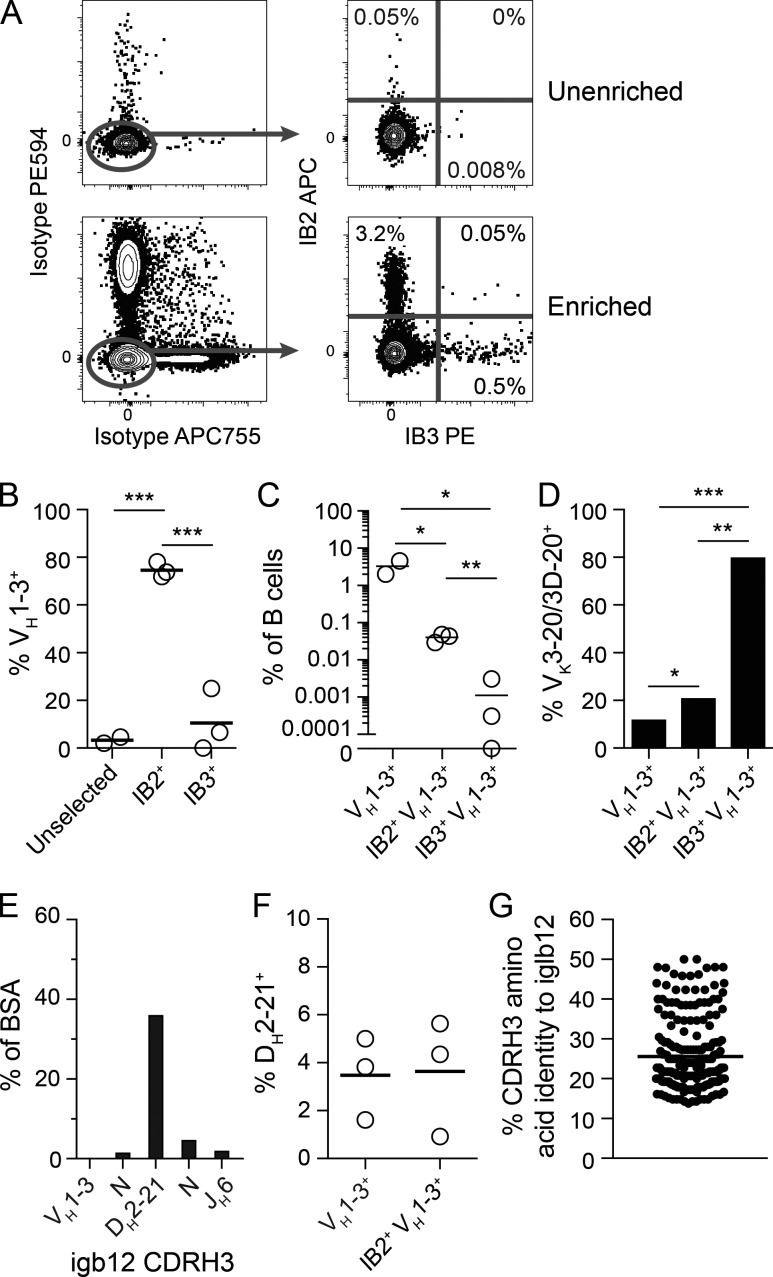
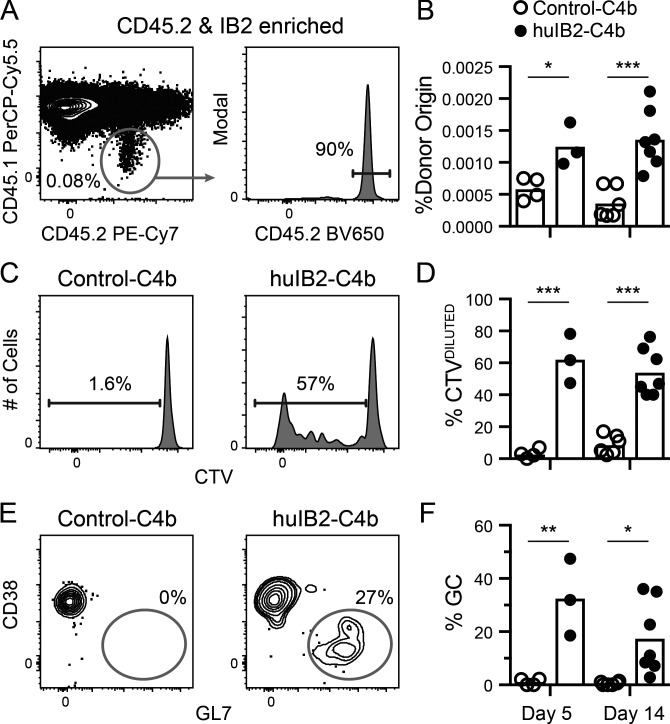
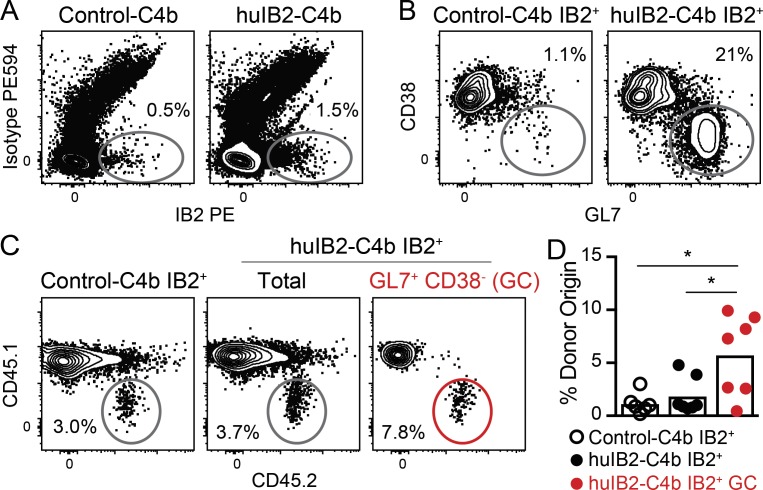
Many tested vaccines fail to provide protection against disease despite the induction of antibodies that bind the pathogen of interest. In light of this, there is much interest in rationally designed subunit vaccines that direct the antibody response to protective epitopes. Here, we produced a panel of anti-idiotype antibodies able to specifically recognize the inferred germline version of the human immunodeficiency virus 1 (HIV-1) broadly neutralizing antibody b12 (iglb12). We determined the crystal structure of two anti-idiotypes in complex with iglb12 and used these anti-idiotypes to identify rare naive human B cells expressing B cell receptors with similarity to iglb12. Immunization with a multimerized version of this anti-idiotype induced the proliferation of transgenic murine B cells expressing the iglb12 heavy chain in vivo, despite the presence of deletion and anergy within this population. Together, our data indicate that anti-idiotypes are a valuable tool for the study and induction of potentially protective antibodies.
© 2019 Bancroft et al.
Figures
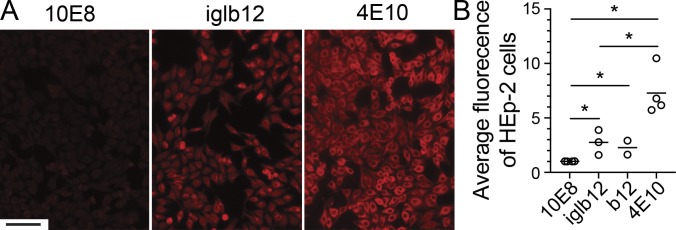
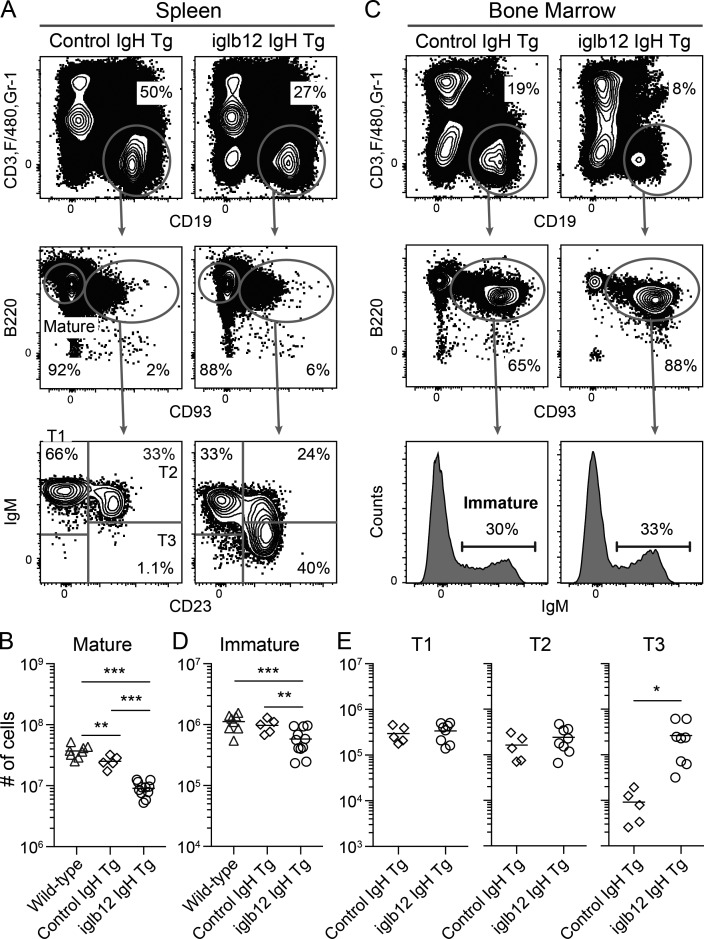
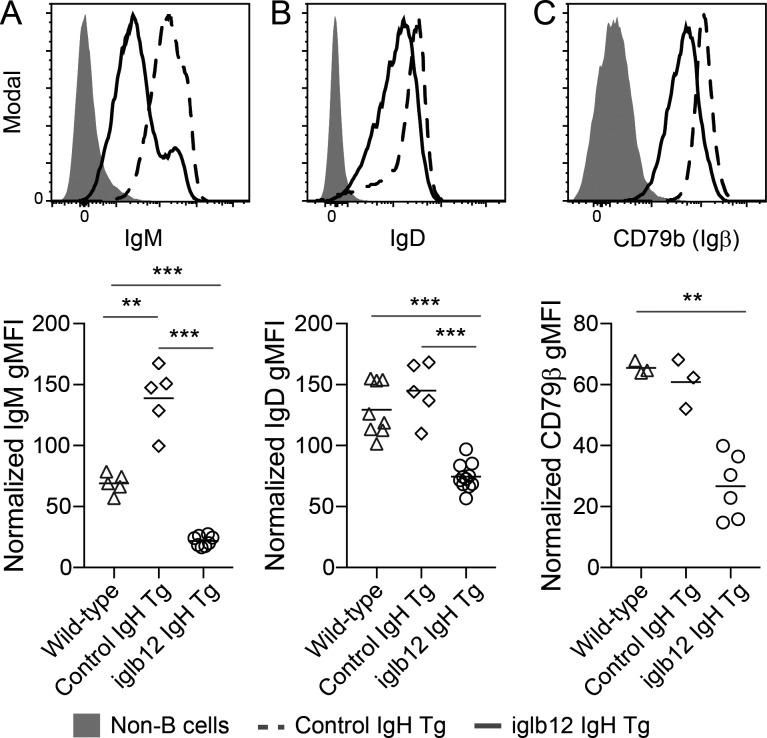
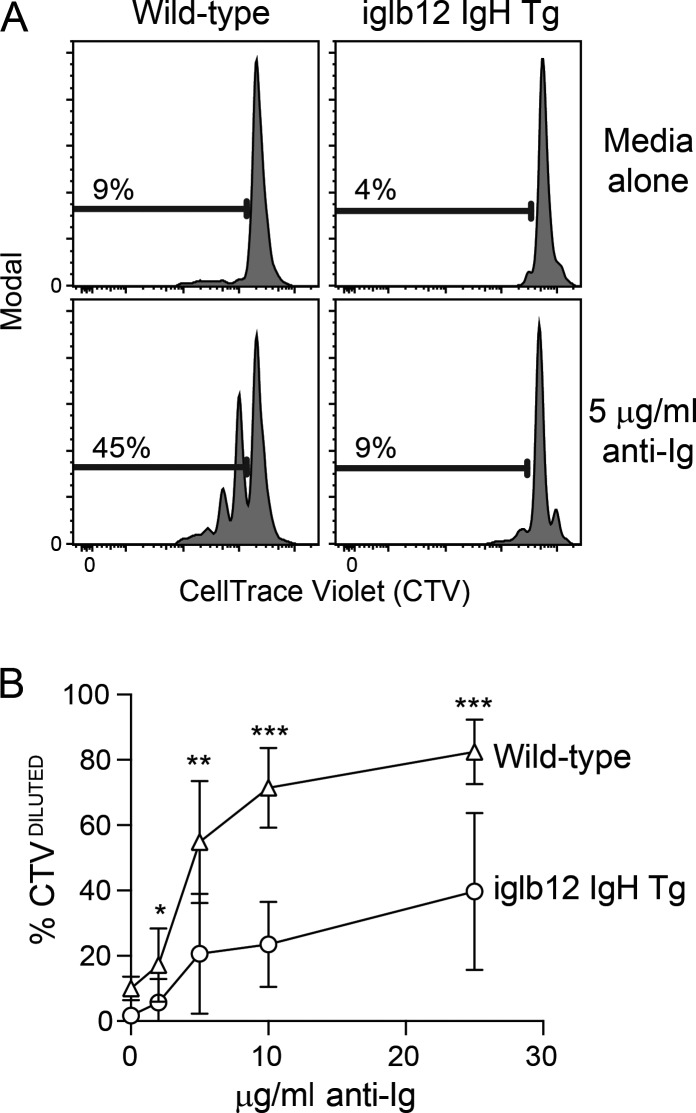
References
-
- Abbott R.K., Lee J.H., Menis S., Skog P., Rossi M., Ota T., Kulp D.W., Bhullar D., Kalyuzhniy O., Havenar-Daughton C., et al. 2018. Precursor Frequency and Affinity Determine B Cell Competitive Fitness in Germinal Centers, Tested with Germline-Targeting HIV Vaccine Immunogens. Immunity. 48:133–146.e6. 10.1016/j.immuni.2017.11.023 - DOI - PMC - PubMed
-
- Adams P.D., Afonine P.V., Bunkóczi G., Chen V.B., Davis I.W., Echols N., Headd J.J., Hung L.W., Kapral G.J., Grosse-Kunstleve R.W., et al. 2010. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 66:213–221. 10.1107/S0907444909052925 - DOI - PMC - PubMed
-
- Agrawal A.S., Ying T., Tao X., Garron T., Algaissi A., Wang Y., Wang L., Peng B.H., Jiang S., Dimitrov D.S., and Tseng C.T.. 2016. Passive Transfer of A Germline-like Neutralizing Human Monoclonal Antibody Protects Transgenic Mice Against Lethal Middle East Respiratory Syndrome Coronavirus Infection. Sci. Rep. 6:31629 10.1038/srep31629 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
