

Multiphase progenetic development shaped the brain of flying archosaurs
- PMID: 31346192
- PMCID: PMC6658547
- DOI: 10.1038/s41598-019-46959-2
Multiphase progenetic development shaped the brain of flying archosaurs
Abstract
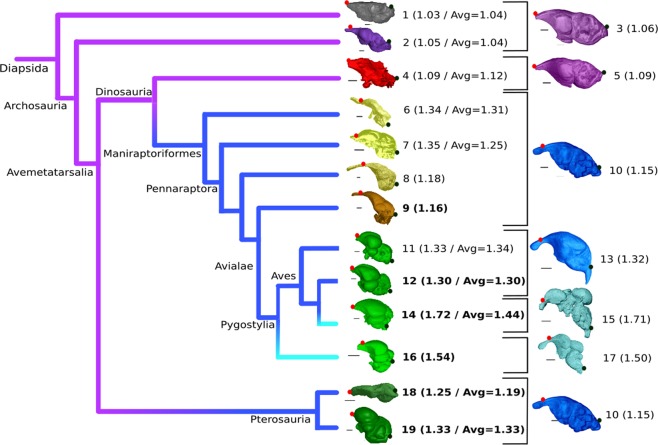
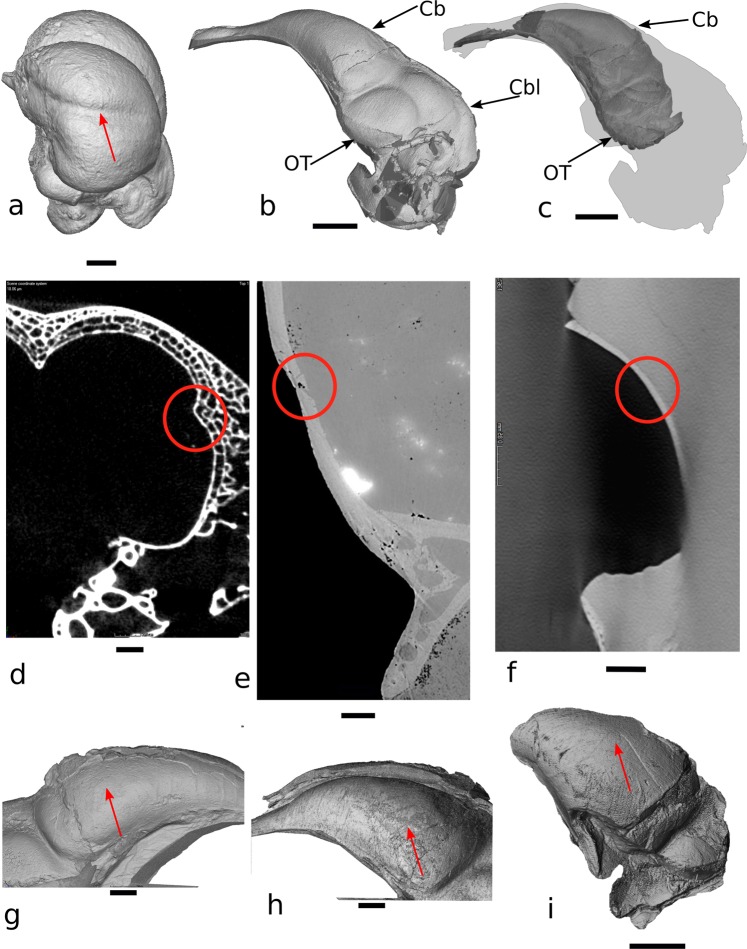
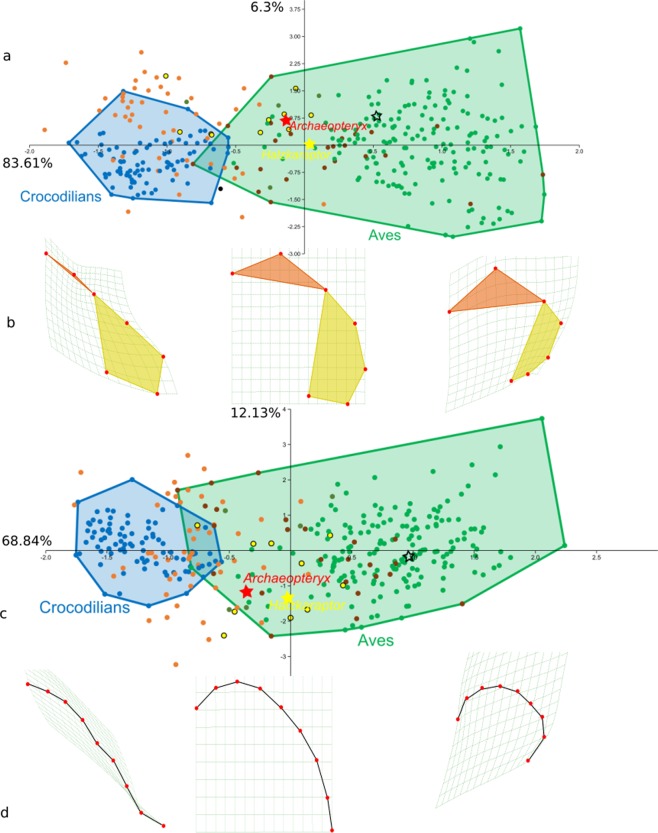
The growing availability of virtual cranial endocasts of extinct and extant vertebrates has fueled the quest for endocranial characters that discriminate between phylogenetic groups and resolve their neural significances. We used geometric morphometrics to compare a phylogenetically and ecologically comprehensive data set of archosaurian endocasts along the deep evolutionary history of modern birds and found that this lineage experienced progressive elevation of encephalisation through several chapters of increased endocranial doming that we demonstrate to result from progenetic developments. Elevated encephalisation associated with progressive size reduction within Maniraptoriformes was secondarily exapted for flight by stem avialans. Within Mesozoic Avialae, endocranial doming increased in at least some Ornithurae, yet remained relatively modest in early Neornithes. During the Paleogene, volant non-neoavian birds retained ancestral levels of endocast doming where a broad neoavian niche diversification experienced heterochronic brain shape radiation, as did non-volant Palaeognathae. We infer comparable developments underlying the establishment of pterosaurian brain shapes.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
