

Dynamic nonlinearities enable direction opponency in Drosophila elementary motion detectors
- PMID: 31346296
- PMCID: PMC6748873
- DOI: 10.1038/s41593-019-0443-y
Dynamic nonlinearities enable direction opponency in Drosophila elementary motion detectors
Abstract
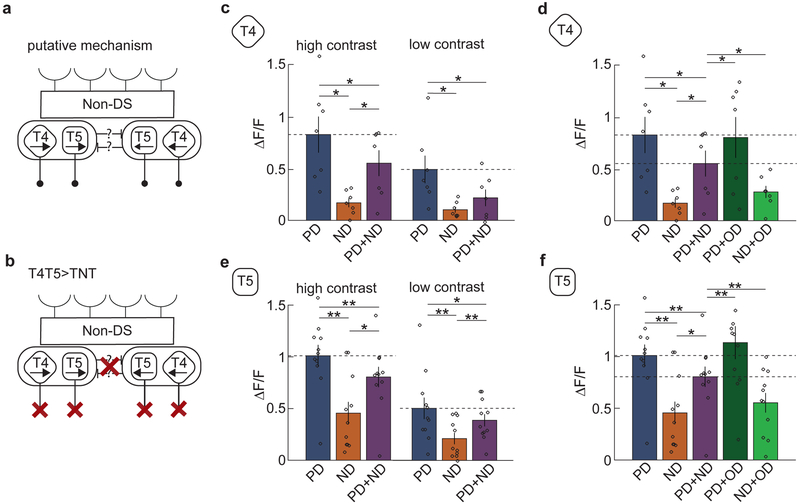
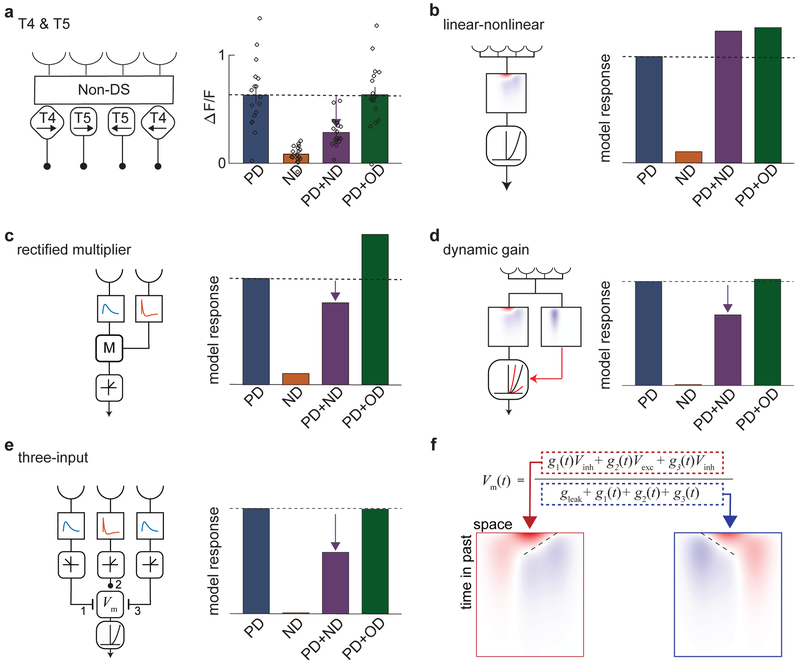
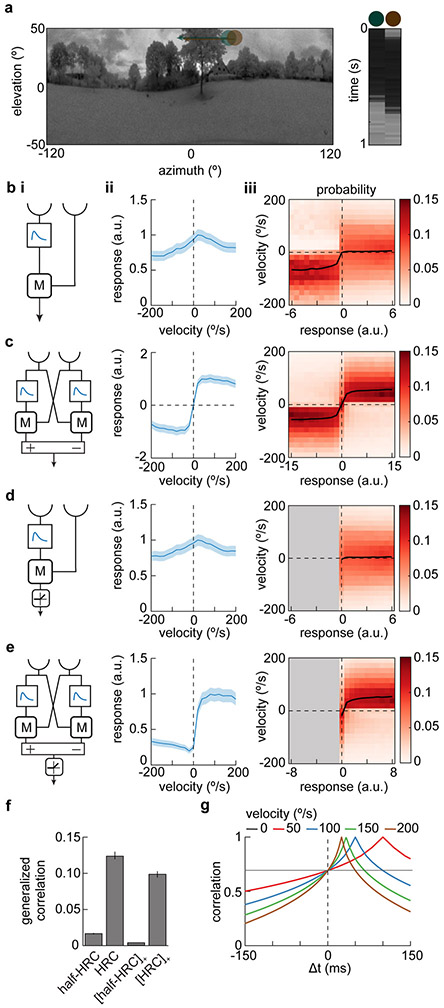
Direction-selective neurons respond to visual motion in a preferred direction. They are direction-opponent if they are also inhibited by motion in the opposite direction. In flies and vertebrates, direction opponency has been observed in second-order direction-selective neurons, which achieve this opponency by subtracting signals from first-order direction-selective cells with opposite directional tunings. Here, we report direction opponency in Drosophila that emerges in first-order direction-selective neurons, the elementary motion detectors T4 and T5. This opponency persists when synaptic output from these cells is blocked, suggesting that it arises from feedforward, not feedback, computations. These observations exclude a broad class of linear-nonlinear models that have been proposed to describe direction-selective computations. However, they are consistent with models that include dynamic nonlinearities. Simulations of opponent models suggest that direction opponency in first-order motion detectors improves motion discriminability by suppressing noise generated by the local structure of natural scenes.
Figures
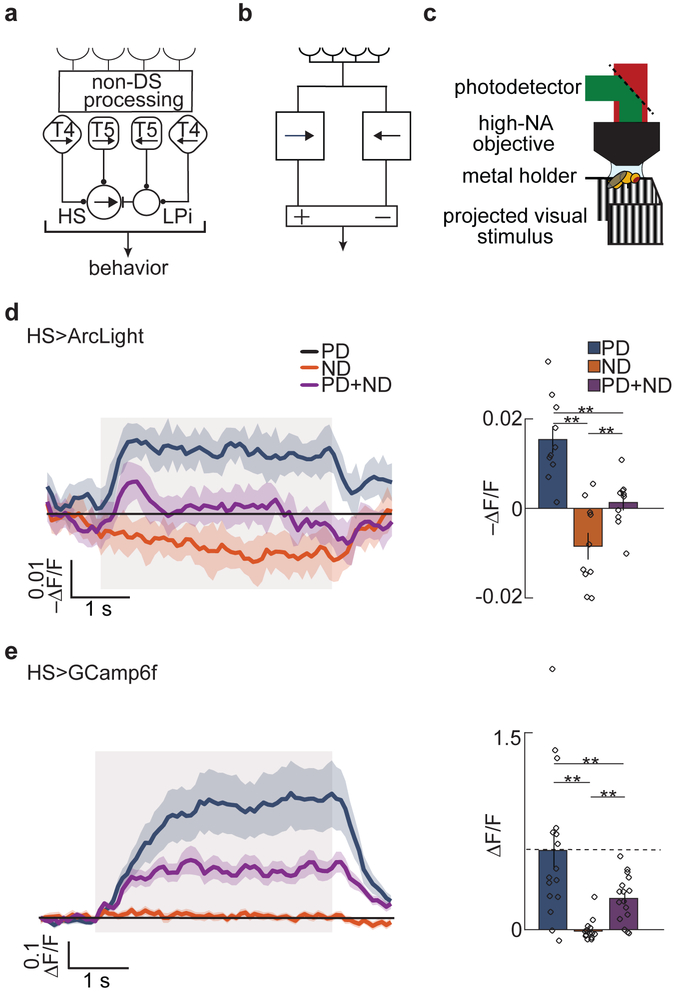
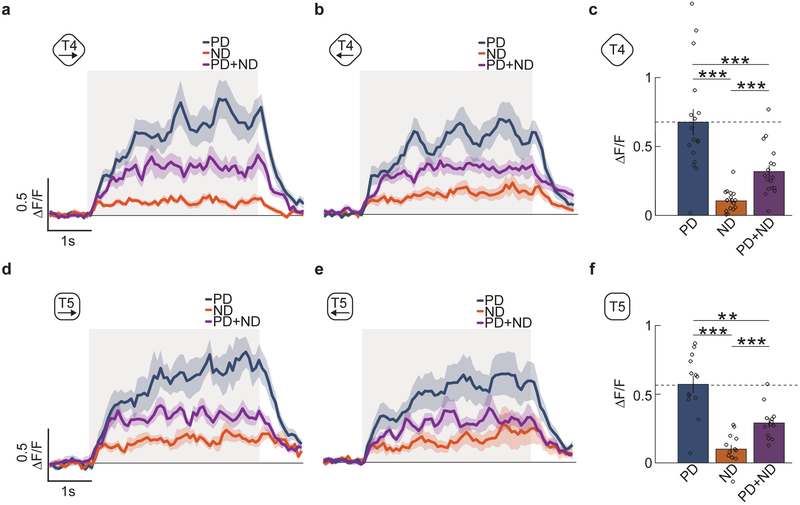
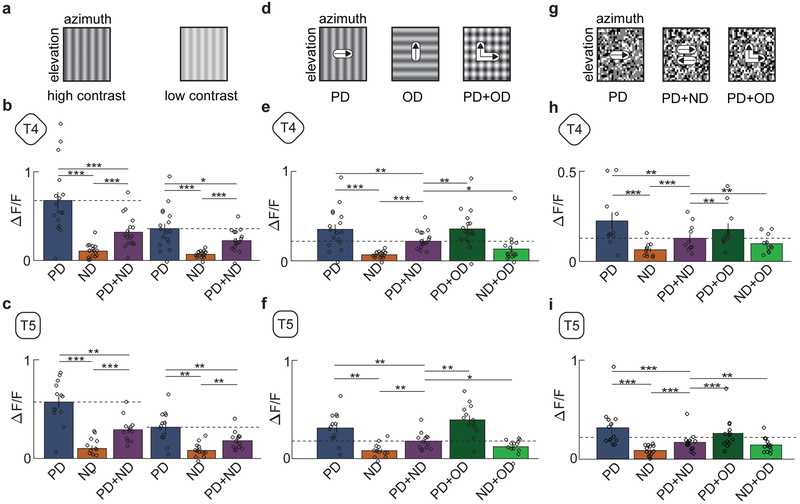
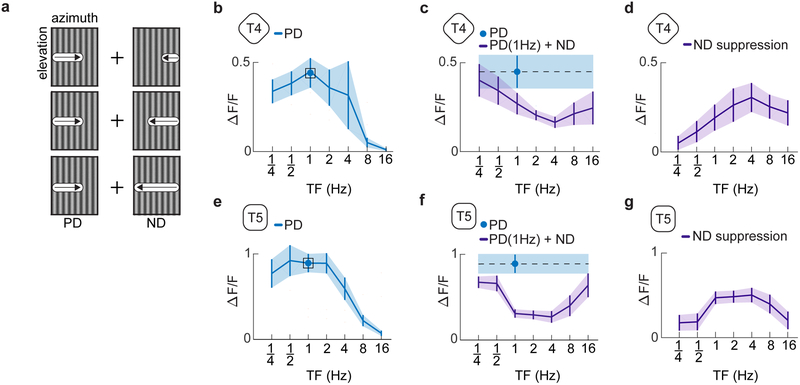
References
Citations
-
- Hausen K Motion sensitive interneurons in the optomotor system of the fly. Biol. Cybern. 45, 143–156 (1982).
-
- Joesch M, Plett J, Borst A & Reiff D Response properties of motion-sensitive visual interneurons in the lobula plate of Drosophila melanogaster. Curr. Biol. 18, 368–374 (2008). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
