

New Insights into Chronological Mobility of Retrotransposons In Vivo
- PMID: 31346359
- PMCID: PMC6617872
- DOI: 10.1155/2019/2818415
New Insights into Chronological Mobility of Retrotransposons In Vivo
Abstract
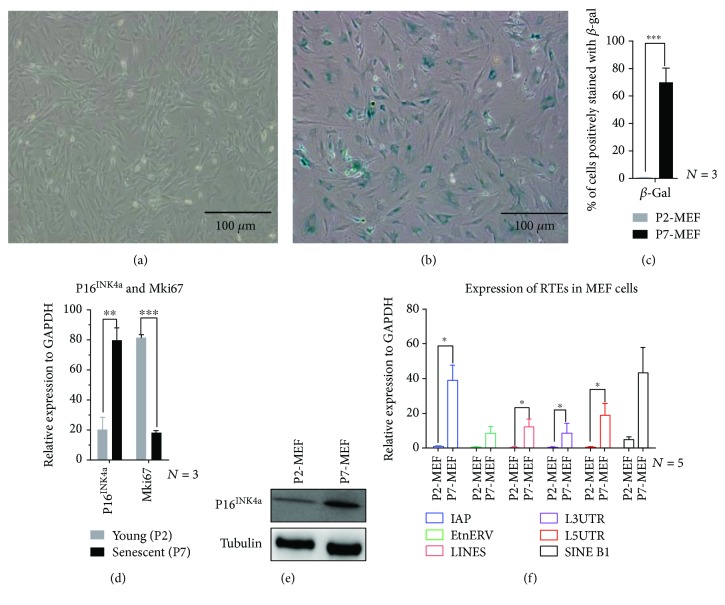
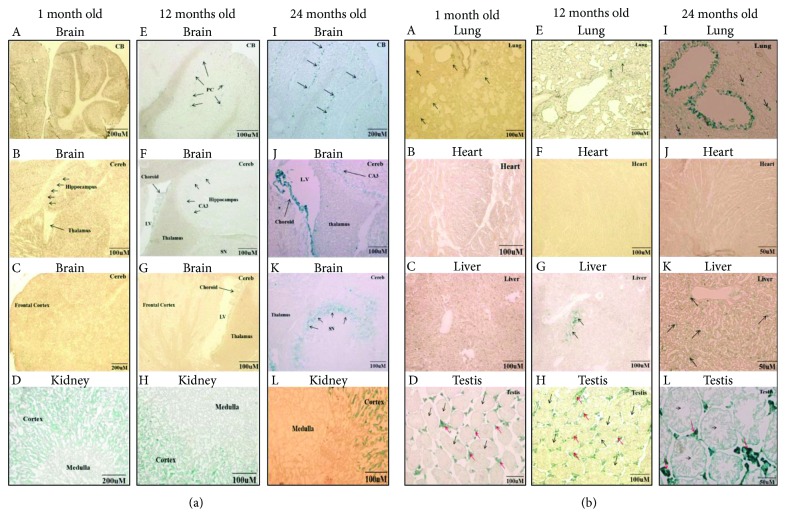
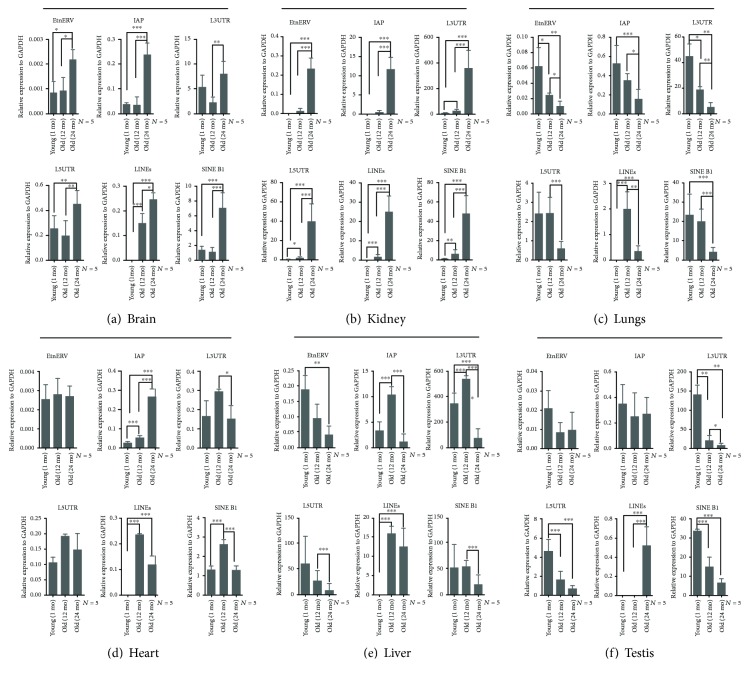
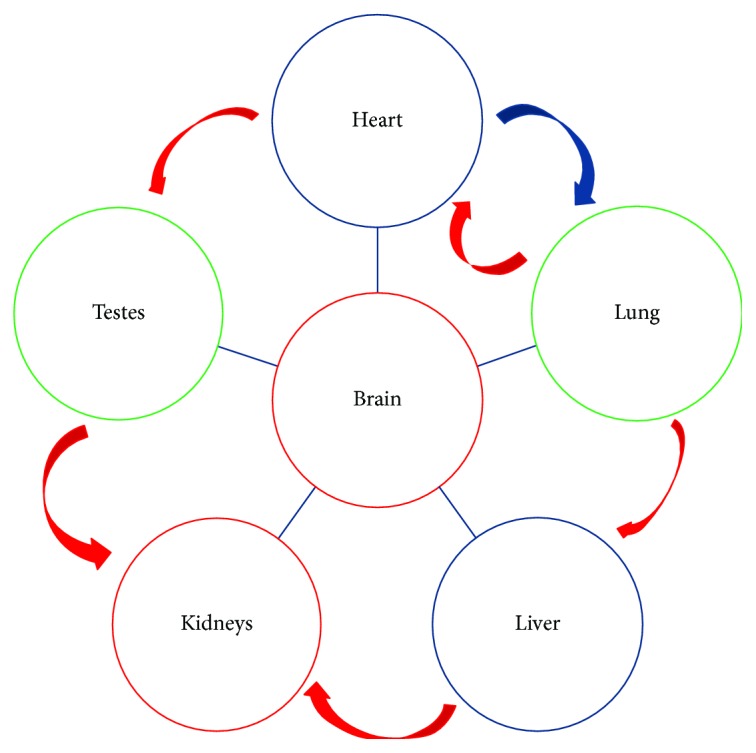
Tissue aging is the gradual decline of physiological homeostasis accompanied with accumulation of senescent cells, decreased clearance of unwanted biological compounds, and depletion of stem cells. Senescent cells were cell cycle arrested in response to various stimuli and identified using distinct phenotypes and changes in gene expression. Senescent cells that accumulate with aging can compromise normal tissue function and inhibit or stop repair and regeneration. Selective removal of senescent cells can slow the aging process and inhibits age-associated diseases leading to extended lifespans in mice and thus provides a possibility for developing antiaging therapy. To monitor the appearance of senescent cells in vivo and target them, a clearer understanding of senescent cell expression markers is needed. We investigated the age-associated expression of three molecular hallmarks of aging: SA-β-gal, P16INK4a, and retrotransposable elements (RTEs), in different mouse tissues during chronological aging. Our data showed that the expression of these markers is variable with aging in the different tissues. P16INK4a showed consistent increases with age in most tissues, while expression of RTEs was variable among different tissues examined. These data suggest that biological changes occurring with physiological aging may be useful in choosing the appropriate timing of therapeutic interventions to slow the aging process or keep more susceptible organs healthier in the aging process.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
