

Population structure of avian malaria parasites
- PMID: 31346436
- PMCID: PMC6635940
- DOI: 10.1002/ece3.5356
Population structure of avian malaria parasites
Abstract
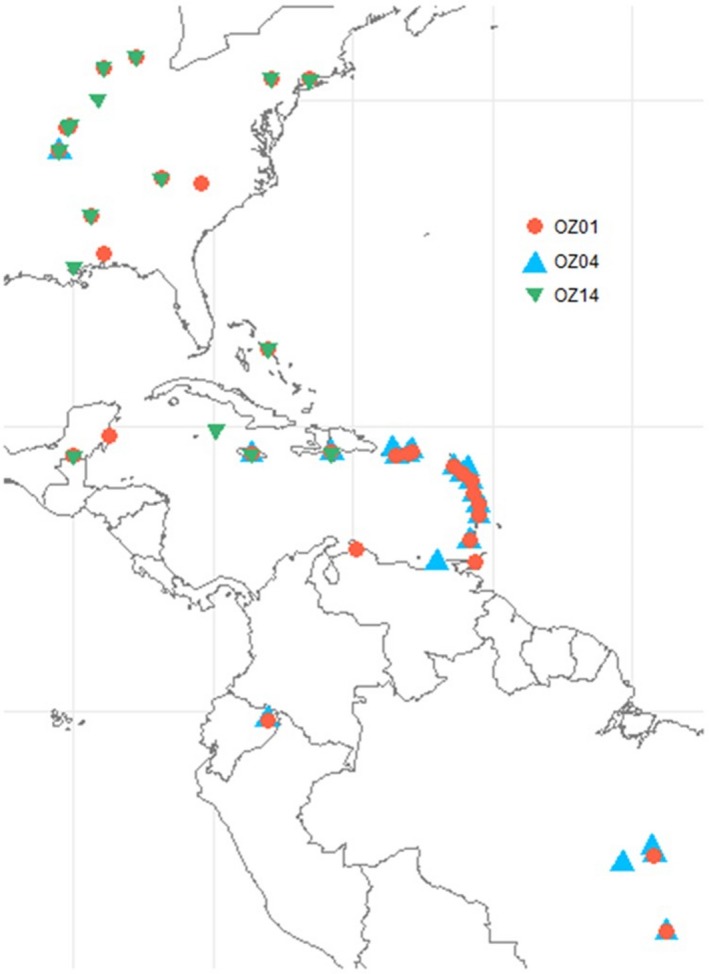
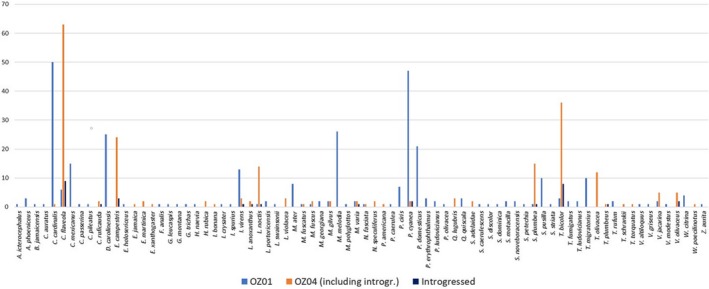
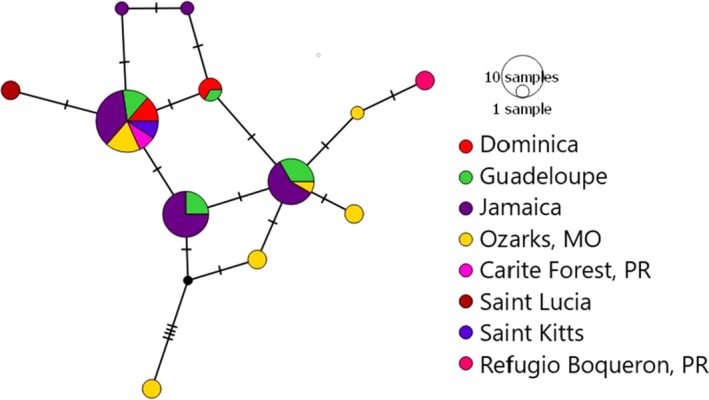
The geographic distribution of genetic diversity in malaria parasite populations (Apicomplexa: Haemosporida) presumably influences local patterns of virulence and the evolution of host-resistance, but little is known about population genetic structure in these parasites. We assess the distribution of genetic diversity in the partial Domain I of apical membrane antigen 1 (AMA1) in three mtDNA-defined lineages of avian Plasmodium to determine spatial population structure and host-parasite genetic relationships. We find that one parasite lineage is genetically differentiated in association with a single host genus and among some locations, but not with respect to other hosts. Two other parasite lineages are undifferentiated with respect to host species but exhibit geographic differentiation that is inconsistent with shared geographic barriers or with isolation-by-distance. Additional differentiation within two other lineages is unassociated with host species or location; in one case, we tentatively interpret this differentiation as the result of mitochondrial introgression from one of the lineages into a second lineage. More sampling of nuclear genetic diversity within populations of avian Plasmodium is needed to rule out coinfection as a possible confounding factor. If coinfections are not responsible for these findings, further assessment is needed to determine the frequency of mitonuclear discordance and its implications for defining parasite lineages based on mitochondrial genetic variation.
Open research badges: This article has earned an Open Data Badge for making publicly available the digitally-shareable data necessary to reproduce the reported results. The data is available at Genbank https://www.ncbi.nlm.nih.gov/genbank/, accession numbers MK965548-MK965653 and MK929797-MK930264.
Keywords: AMA1; Haemoproteus; Haemosporida; Plasmodium; avian malaria; host–parasite genetic relationship; introgression; spatial population structure.
Conflict of interest statement
All authors declare no conflict of interest.
Figures







References
-
- Abouie Mehrizi, A. , Sepehri, M. , Karimi, F. , Djadid, N. D. , & Zakeri, S. (2013). Infection, genetics and evolution population genetics, sequence diversity and selection in the gene encoding the Plasmodium falciparum apical membrane antigen 1 in clinical isolates from the South‐East of Iran. Infection, Genetics and Evolution, 17, 51–61. 10.1016/j.meegid.2013.03.042 - DOI - PubMed
-
- Arnott, A. , Wapling, J. , Mueller, I. , Ramsland, P. A. , Siba, P. M. , Reeder, J. C. , & Barry, A. (2014). Distinct Patterns of Diversity, Population Structure and Evolution in the AMA1 Genes of Sympatric Plasmodium falciparum and Plasmodium vivax Populations of Papua New Guinea from an Area of Similarly High Transmission. Malaria Journal, 13(233), 1–16. - PMC - PubMed
-
- Bai, T. , Becker, M. , Gupta, A. , Strike, P. , Murphy, V. J. , Anders, R. F. , & Batchelor, A. H. (2005). Structure of AMA1 from Plasmodium falciparum Reveals a Clustering of Polymorphisms That Surround a Conserved Hydrophobic Pocket. Proceedings of the National Academy of Science, 102, 12736–12741. - PMC - PubMed
-
- Bell, J. R. (2008). A simple way to treat PCR products prior to sequencing using ExoSAP‐IT ®. BioTechniques, 44(6), 2008. - PubMed
-
- Bensch, S. , Hellgren, O. , & Pérez‐Tris, J. (2009). MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Molecular Ecology Resources, 9(5), 1353–1358. - PubMed
LinkOut - more resources
Full Text Sources
