

Origin and evolution of the major histocompatibility complex class I region in eutherian mammals
- PMID: 31346446
- PMCID: PMC6636196
- DOI: 10.1002/ece3.5373
Origin and evolution of the major histocompatibility complex class I region in eutherian mammals
Abstract
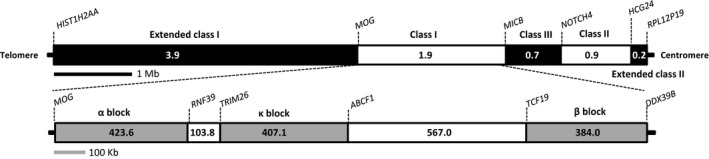
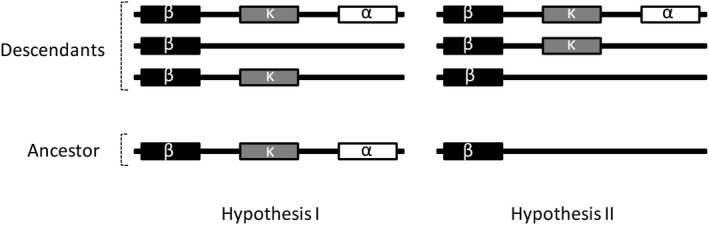
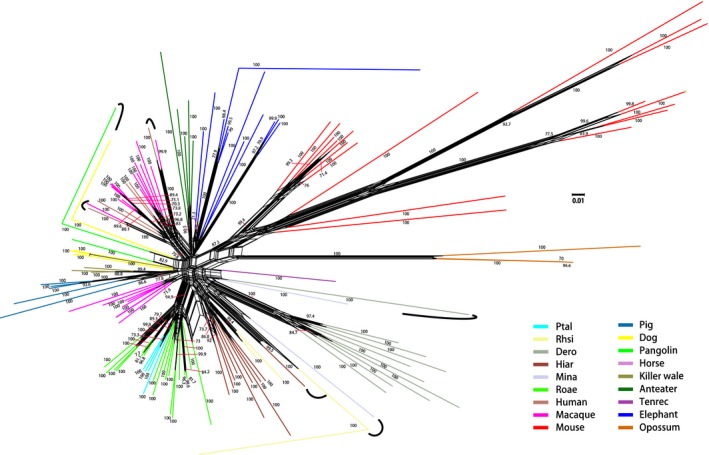
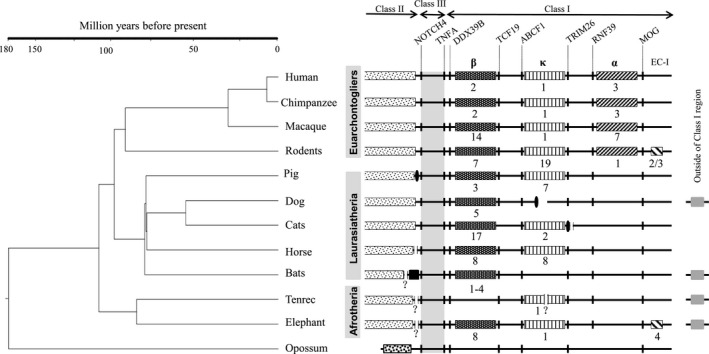
Major histocompatibility complex (MHC) genes in vertebrates are vital in defending against pathogenic infections. To gain new insights into the evolution of MHC Class I (MHCI) genes and test competing hypotheses on the origin of the MHCI region in eutherian mammals, we studied available genome assemblies of nine species in Afrotheria, Xenarthra, and Laurasiatheria, and successfully characterized the MHCI region in six species. The following numbers of putatively functional genes were detected: in the elephant, four, one, and eight in the extended class I region, and κ and β duplication blocks, respectively; in the tenrec, one in the κ duplication block; and in the four bat species, one or two in the β duplication block. Our results indicate that MHCI genes in the κ and β duplication blocks may have originated in the common ancestor of eutherian mammals. In the elephant, tenrec, and all four bats, some MHCI genes occurred outside the MHCI region, suggesting that eutherians may have a more complex MHCI genomic organization than previously thought. Bat-specific three- or five-amino-acid insertions were detected in the MHCI α1 domain in all four bats studied, suggesting that pathogen defense in bats relies on MHCIs having a wider peptide-binding groove, as previously assayed by a bat MHCI gene with a three-amino-acid insertion showing a larger peptide repertoire than in other mammals. Our study adds to knowledge on the diversity of eutherian MHCI genes, which may have been shaped in a taxon-specific manner.
Keywords: MHC; bats; comparative genomics; evolution; mammals; origin.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


References
-
- Abduriyim, S. , Nishita, Y. , Kosintsev, P. A. , Raichev, E. , Väinölä, R. , Kryukov, A. P. , … Masuda, R. (2017). Diversity and evolution of MHC class II DRB gene in the Eurasian badger genus Meles (Mammalia: Mustelidae). Biological Journal of the Linnean Society, 122(2), 258–273. 10.1093/biolinnean/blx077 - DOI
LinkOut - more resources
Full Text Sources
Research Materials
