

COX2 is induced in the ovarian epithelium during ovulatory wound repair and promotes cell survival†
- PMID: 31347667
- PMCID: PMC6877777
- DOI: 10.1093/biolre/ioz134
COX2 is induced in the ovarian epithelium during ovulatory wound repair and promotes cell survival†
Abstract
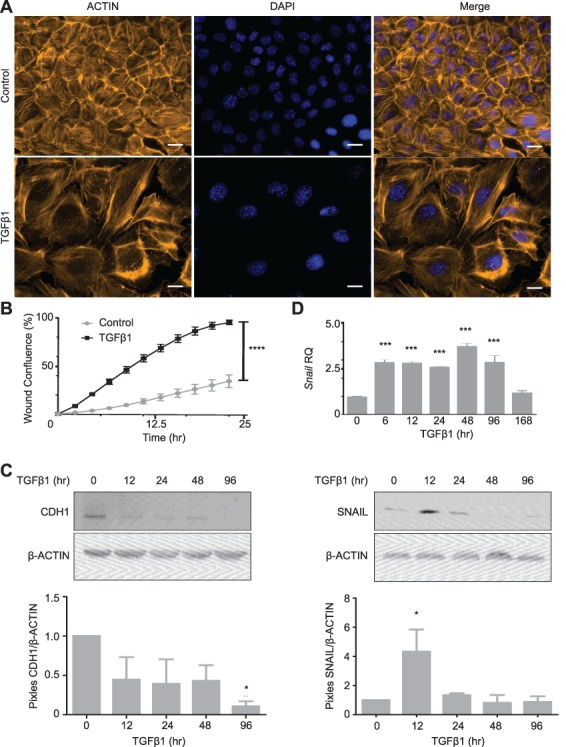
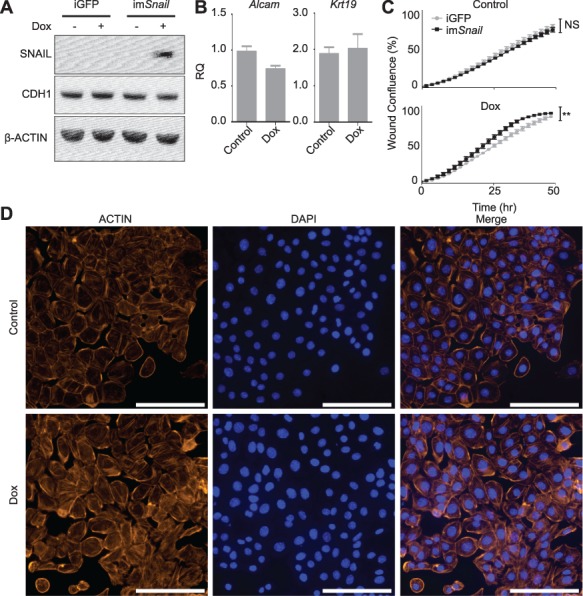
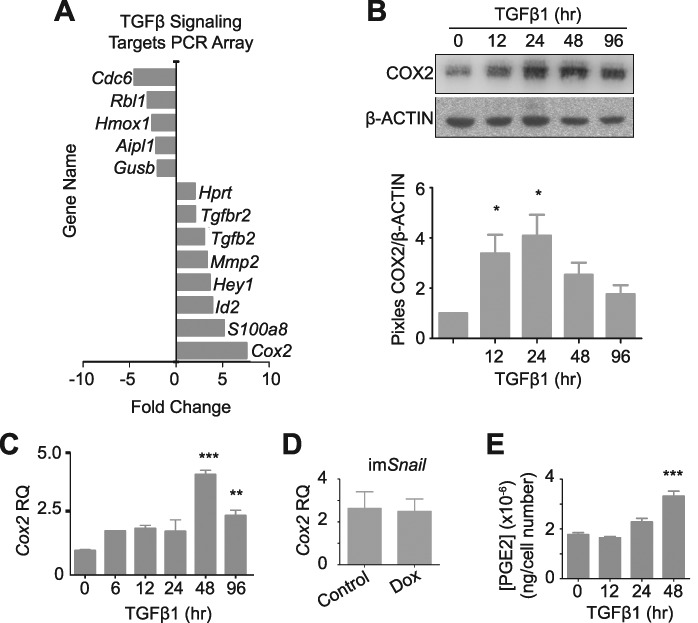
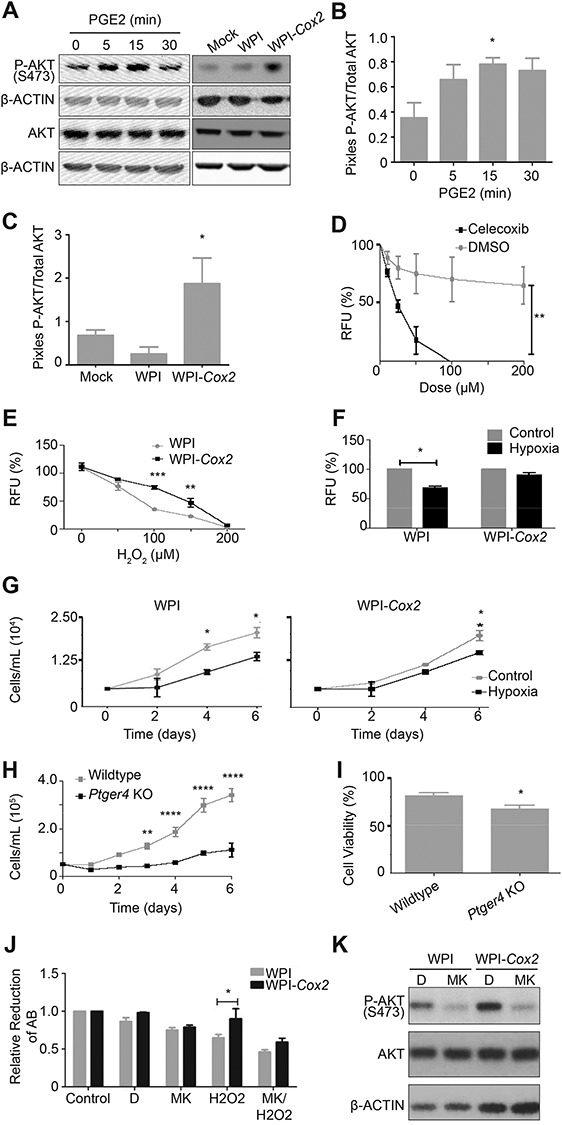
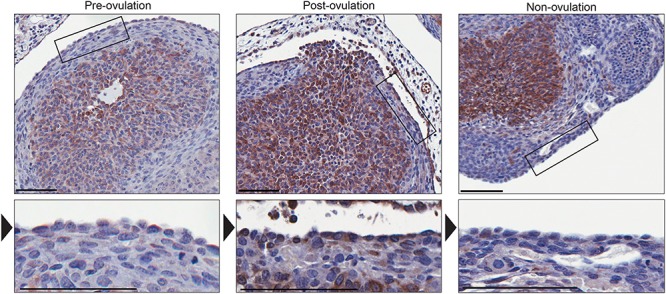
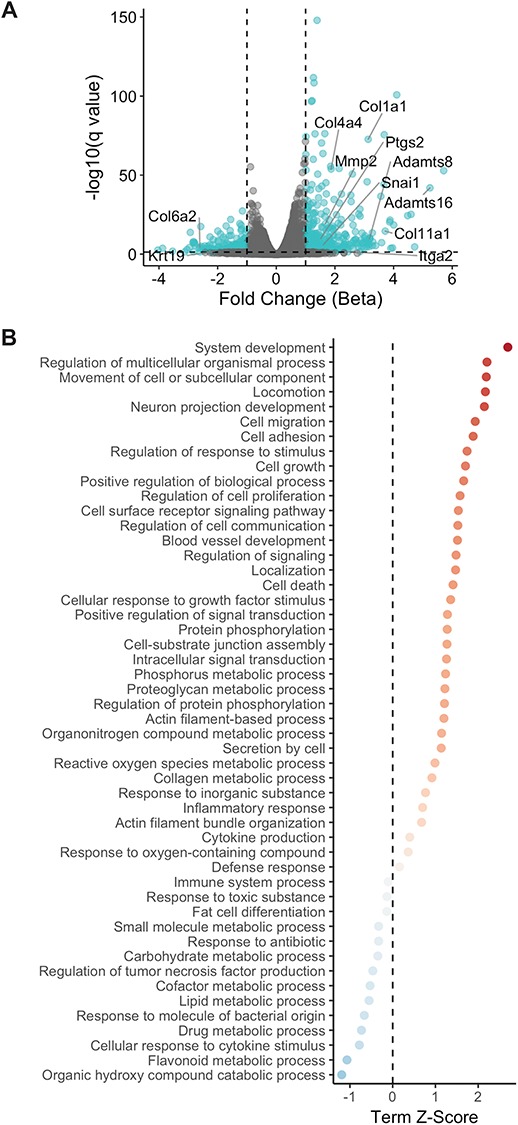
The ovarian surface epithelium (OSE) is a monolayer of cells surrounding the ovary that is ruptured during ovulation. After ovulation, the wound is repaired, however, this process is poorly understood. In epithelial tissues, wound repair is mediated by an epithelial-to-mesenchymal transition (EMT). Transforming Growth Factor Beta-1 (TGFβ1) is a cytokine commonly known to induce an EMT and is present throughout the ovarian microenvironment. We, therefore, hypothesized that TGFβ1 induces an EMT in OSE cells and activates signaling pathways important for wound repair. Treating primary cultures of mouse OSE cells with TGFβ1 induced an EMT mediated by TGFβRI signaling. The transcription factor Snail was the only EMT-associated transcription factor increased by TGFβ1 and, when overexpressed, was shown to increase OSE cell migration. A polymerase chain reaction array of TGFβ signaling targets determined Cyclooxygenase-2 (Cox2) to be most highly induced by TGFβ1. Constitutive Cox2 expression modestly increased migration and robustly enhanced cell survival, under stress conditions similar to those observed during wound repair. The increase in Snail and Cox2 expression with TGFβ1 was reproduced in human OSE cultures, suggesting these responses are conserved between mouse and human. Finally, the induction of Cox2 expression in OSE cells during ovulatory wound repair was shown in vivo, suggesting TGFβ1 increases Cox2 to promote wound repair by enhancing cell survival. These data support that TGFβ1 promotes ovulatory wound repair by induction of an EMT and activation of a COX2-mediated pro-survival pathway. Understanding ovulatory wound repair may give insight into why ovulation is the primary non-hereditary risk factor for ovarian cancer.
Keywords: Cyclooxygenase 2; epithelial-to-mesenchymal transition; ovarian surface epithelium; transforming growth factor beta 1; wound repair.
© The Author(s) 2019. Published by Oxford University Press on behalf of Society for the Study of Reproduction. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Lim H, Paria BC, Das SK, Dinchuk JE, Langenbach R, Trzaskos JM, Dey SK. Multiple female reproductive failures in cyclooxygenase 2–deficient mice. Cell 1997; 91:197–208. - PubMed
-
- Richards JS. Ovulation: New factors that prepare the oocyte for fertilization. Mol Cell Endocrinol 2005; 234:75–79. - PubMed
-
- Murdoch WJ. Ovarian surface epithelium during ovulatory and anovulatory ovine estrous cycles. Anat Rec 1994; 240:322–326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
