

Differences in Steap3 expression are a mechanism of genetic variation of RBC storage and oxidative damage in mice
- PMID: 31350307
- PMCID: PMC6693009
- DOI: 10.1182/bloodadvances.2019000605
Differences in Steap3 expression are a mechanism of genetic variation of RBC storage and oxidative damage in mice
Abstract
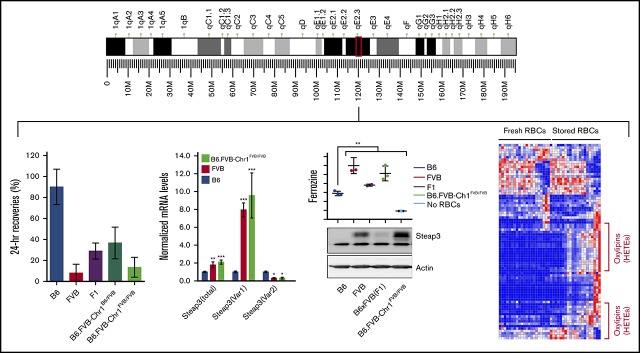
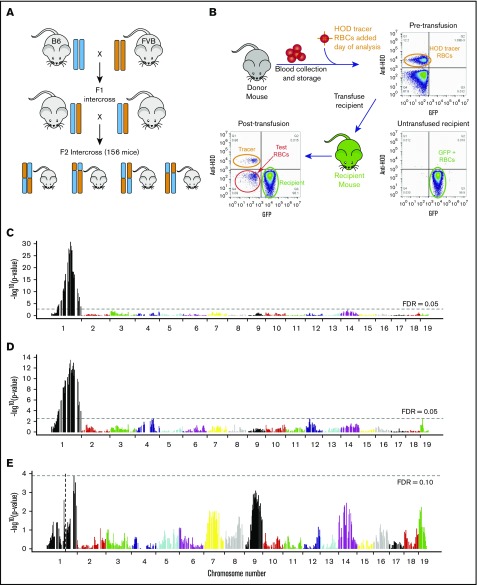
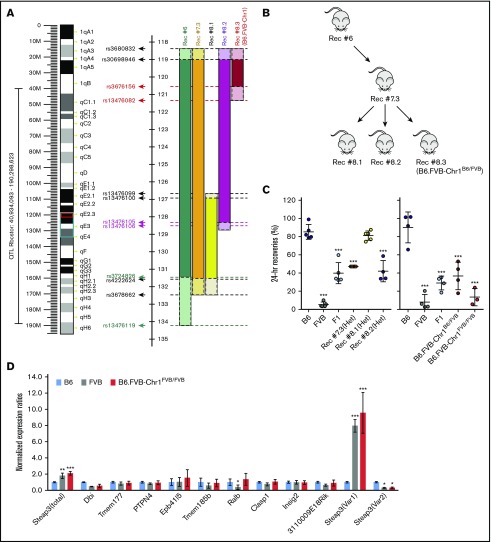
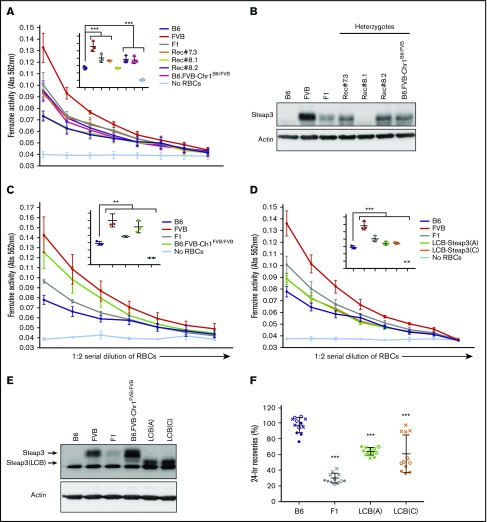
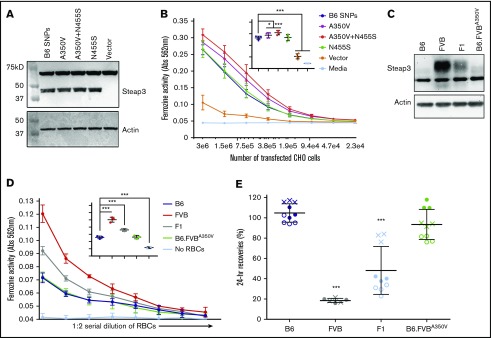
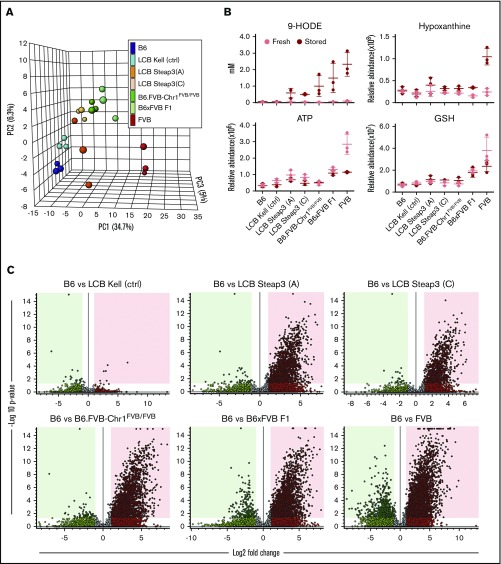
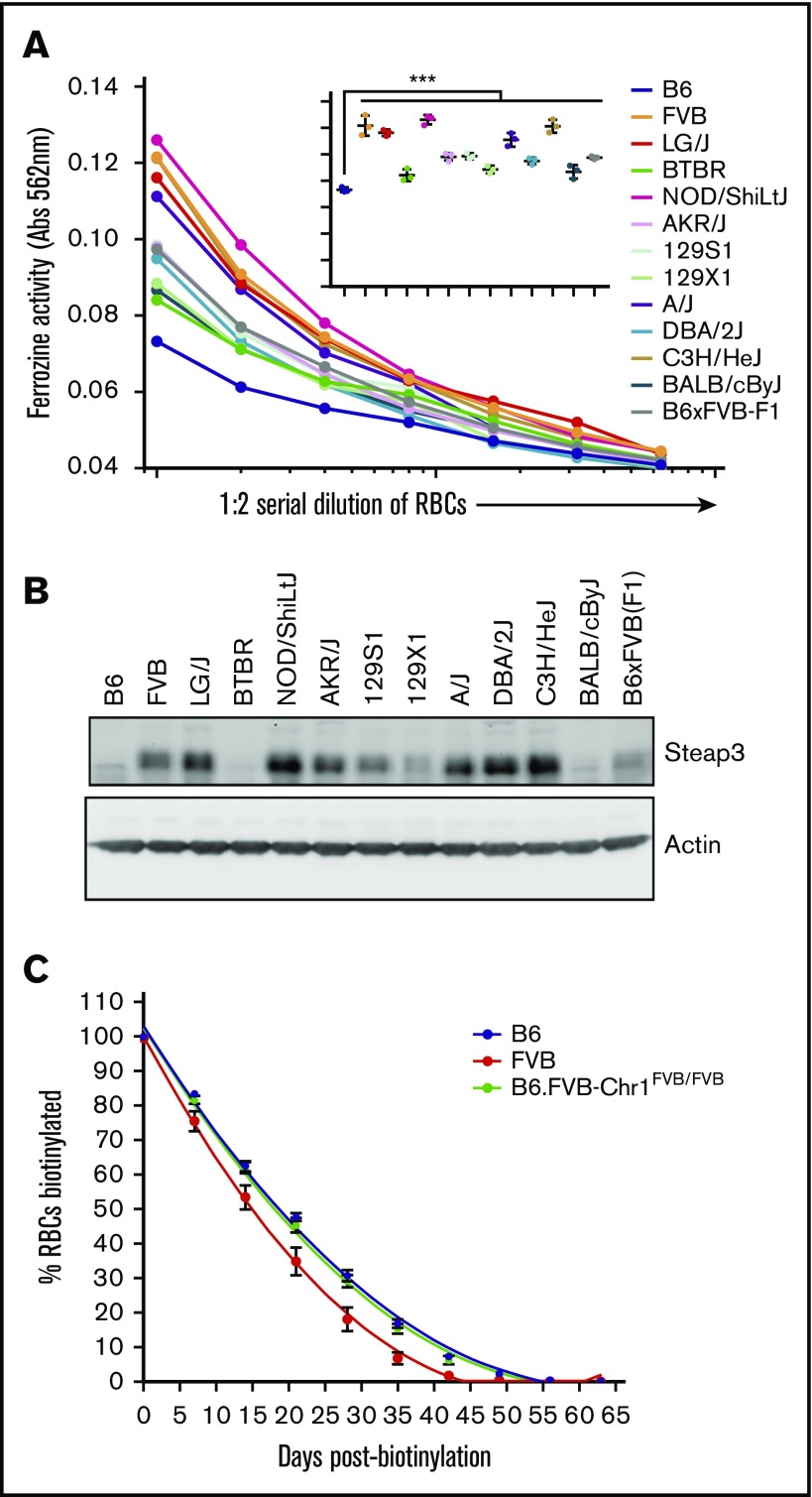
Red blood cells (RBCs) are the most numerous cell type in the body and serve a vital purpose of delivering oxygen to essentially all tissues. In addition to the central role of RBCs in health and disease, RBC storage is a requirement for the >90 million units of RBC transfusions given to millions of recipients each year, worldwide. It is well known that there is genetic donor-to-donor variability in how human RBCs store, rendering blood a nonstandardized therapeutic with a wide range of biological properties from unit to unit, by the time it is transfused. As with humans, genetic variation exists in how murine RBCs, from different strains of mice, store and perform after transfusion. The genetic mechanisms for variation, in humans and mice, both remain obscure. Combining advanced metabolomics, genetics, and molecular and cellular biology approaches, we identify genetic variation in six-transmembrane epithelial antigen of prostate 3 (Steap3) expression as a critical and previously unrecognized mechanism of oxidative damage of RBCs during storage. Increased levels of Steap3 result in degradation of cellular membrane through lipid peroxidation, leading to failure of RBC homeostasis and hemolysis/clearance of RBCs. This article is the first report of a role of Steap3 in mature RBCs; it defines a new mechanism of redox biology in RBCs with a substantial effect upon RBC function and provides a novel mechanistic determinant of genetic variation of RBC storage.
© 2019 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: Bloodworks NW has filed a patent on using Steap3 as a predictor of RBC storage. J.C.Z. and K.d.W. are inventors on this application (US20170276665A1). The remaining authors declare no competing financial interests.
Figures
References
-
- Dumont LJ, AuBuchon JP. Evaluation of proposed FDA criteria for the evaluation of radiolabeled red cell recovery trials. Transfusion. 2008;48(6):1053-1060. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
