

Priming of Plant Resistance to Heat Stress and Tomato Yellow Leaf Curl Thailand Virus With Plant-Derived Materials
- PMID: 31354773
- PMCID: PMC6640737
- DOI: 10.3389/fpls.2019.00906
Priming of Plant Resistance to Heat Stress and Tomato Yellow Leaf Curl Thailand Virus With Plant-Derived Materials
Abstract
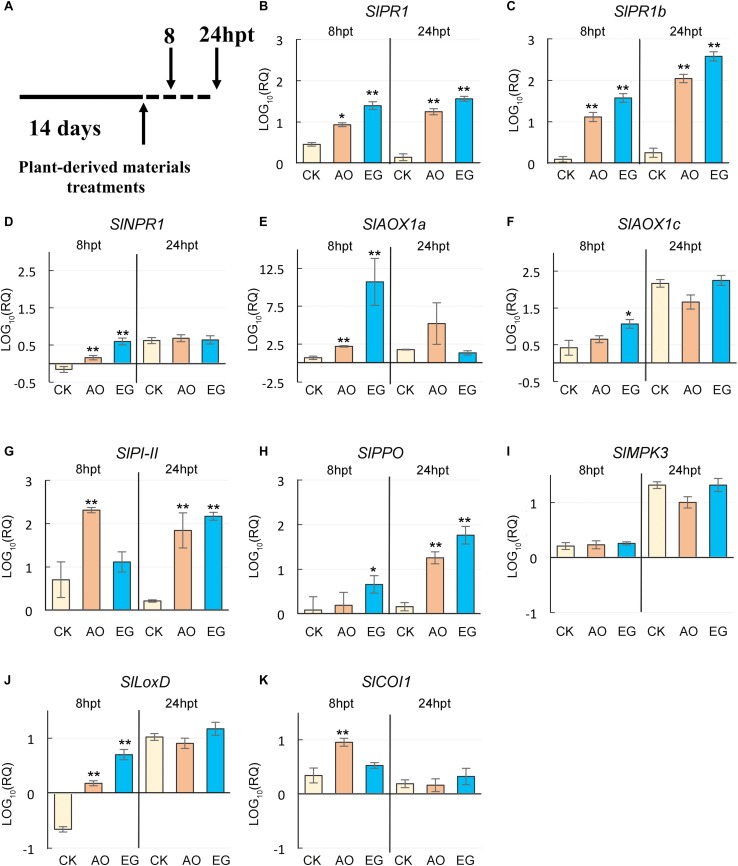
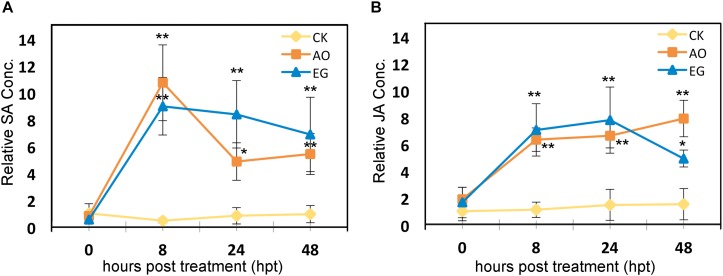
Plants are often simultaneously exposed to diverse environmental stresses, and can tune suitable responses to them through hormones. Salicylic acid (SA) and jasmonic acid (JA) signaling pathways are known to enhance resistance against heat stress and tomato yellow leaf curl Thailand virus (TYLCTHV) infection. However, there is limited information regarding alternative natural priming agents against heat stress and viruses. In this study, two plant-derived priming agents, eugenol and anise oil, were tested for their roles in conferring thermotolerance and virus resistance in tomato plants. Under heat stress, the survival rates and average fresh weight were higher in plants treated with eugenol or anise oil than in control plants. These two priming agents were further tested for antiviral activities. After TYLCTHV infection, the disease incidence and relative abundance of TYLCTHV were lower in anise oil- and eugenol-treated plants than in control plants. Further analyses revealed that a few SA, JA, and RNA silencing genes were enhanced in the former. Moreover, SA, JA, and H2O2 contents increased considerably after eugenol and anise oil treatments. Our findings imply that anise oil and eugenol initiated SA- and JA-mediated defenses to promote thermotolerance and antiviral responses of tomato plants.
Keywords: anise oil; eugenol; jasmonic acid; salicylic acid; thermotolerance; tomato (Solanum lycopersicum); tomato yellow leaf curl Thailand virus (TYLCTHV).
Figures






References
LinkOut - more resources
Full Text Sources