

Inhibition of osteoclastogenesis by opsonized Porphyromonas gingivalis
- PMID: 31355360
- PMCID: PMC6660169
- DOI: 10.1096/fba.2018-00018
Inhibition of osteoclastogenesis by opsonized Porphyromonas gingivalis
Abstract
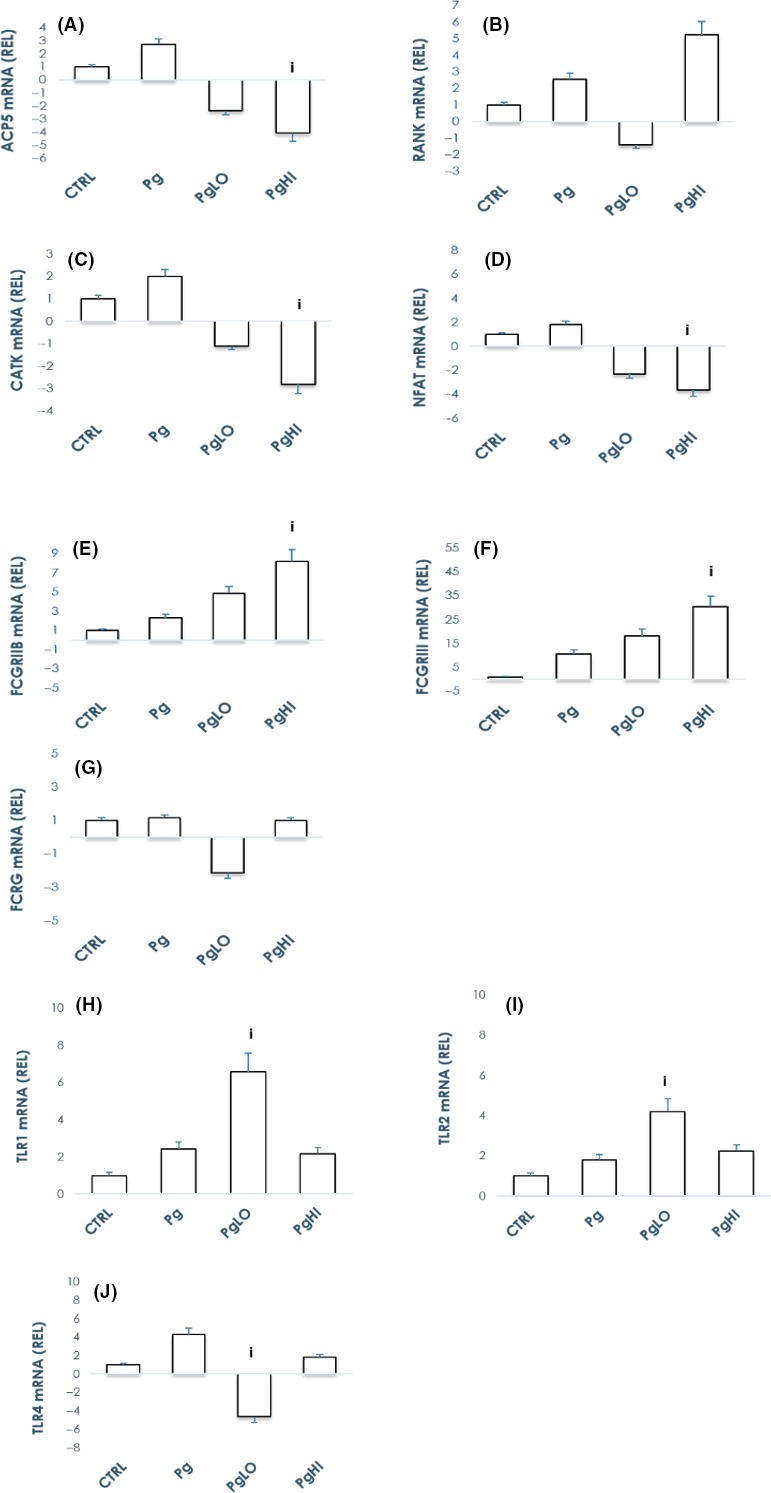
A crucial step in the pathogenesis of periodontal disease (PD) is activation of osteoclasts (OC) by numerous virulence factors produced by Porphyromonas gingivalis (Pg). To understand pathogenesis of periodontal disease and the role of specific adaptive immune responses, effects of antibodies on Pg-induced OC differentiation and function were investigated. Human peripheral blood-derived monocytes were differentiated in vitro to osteoclasts in the presence or absence of: a) Pg; b) antibodies to Pg; and c) antibody-opsonized Pg. Findings suggest significant induction of osteoclastogenesis by Pg when compared to control cultures, whereas opsonization decreased osteoclastogenesis by 45%. Immune receptor gene expression profile in the presence of opsonized Pg showed marked up-regulation of TLR1 (3-fold) and TLR2 (2-fold) along with FcγRIIB (2-fold) and FcγRIII receptors (5-fold), but not TLR4 and FcRγ receptors. Interestingly, blocking FcγRIIB, but not FcγRIII receptor, reversed the inhibitory effects of opsonized Pg suggesting a critical role played by FcγRIIB in osteoclastogenesis. Furthermore, opsonized Pg transformed OC precursors to a "macrophage phenotype" suggesting a bone protective role of the immune complexes in modulating osteoclastogenesis, probably by competing as an agonist for PRRs, and inducing selective activation of FcγRs with simultaneous suppression of FcRγ which regulates bone resorptive process. Further defining effective antibody isotypes, avidity, and antigenic specificity could improve targets for eliciting protective immunity.
Keywords: bone loss; immune response; macrophages; osteoclast; periodontitis.
Conflict of interest statement
Conflict of Interest Statement: Authors report no conflicts of interest related to this study.
Figures




Similar articles
-
A challenge with Porphyromonas gingivalis differentially affects the osteoclastogenesis potential of periodontal ligament fibroblasts from periodontitis patients and non-periodontitis donors.J Clin Periodontol. 2014 Feb;41(2):95-103. doi: 10.1111/jcpe.12186. Epub 2013 Nov 19. J Clin Periodontol. 2014. PMID: 24164598
-
Phosphoglycerol dihydroceramide, a distinctive ceramide produced by Porphyromonas gingivalis, promotes RANKL-induced osteoclastogenesis by acting on non-muscle myosin II-A (Myh9), an osteoclast cell fusion regulatory factor.Biochim Biophys Acta Mol Cell Biol Lipids. 2017 May;1862(5):452-462. doi: 10.1016/j.bbalip.2017.01.008. Epub 2017 Jan 31. Biochim Biophys Acta Mol Cell Biol Lipids. 2017. PMID: 28153611 Free PMC article.
-
Micromolar sodium fluoride mediates anti-osteoclastogenesis in Porphyromonas gingivalis-induced alveolar bone loss.Int J Oral Sci. 2015 Dec 18;7(4):242-9. doi: 10.1038/ijos.2015.28. Int J Oral Sci. 2015. PMID: 26674426 Free PMC article.
-
Immune response: the key to bone resorption in periodontal disease.J Periodontol. 2005 Nov;76(11 Suppl):2033-41. doi: 10.1902/jop.2005.76.11-S.2033. J Periodontol. 2005. PMID: 16277573 Review.
-
Are Sphingolipids and Serine Dipeptide Lipids Underestimated Virulence Factors of Porphyromonas gingivalis?Infect Immun. 2018 Jun 21;86(7):e00035-18. doi: 10.1128/IAI.00035-18. Print 2018 Jul. Infect Immun. 2018. PMID: 29632248 Free PMC article. Review.
Cited by
-
Statins Modulate Microenvironmental Cues Driving Macrophage Polarization in Simulated Periodontal Inflammation.Cells. 2023 Jul 29;12(15):1961. doi: 10.3390/cells12151961. Cells. 2023. PMID: 37566040 Free PMC article.
-
Age and Periodontal Health - Immunological View.Curr Oral Health Rep. 2018 Dec;5(4):229-241. doi: 10.1007/s40496-018-0202-2. Epub 2018 Nov 7. Curr Oral Health Rep. 2018. PMID: 30555774 Free PMC article.
-
The current landscape of microRNAs (miRNAs) in bacterial pneumonia: opportunities and challenges.Cell Mol Biol Lett. 2022 Aug 19;27(1):70. doi: 10.1186/s11658-022-00368-y. Cell Mol Biol Lett. 2022. PMID: 35986232 Free PMC article. Review.
-
Oral microbiome interactions with gingival gene expression patterns for apoptosis, autophagy and hypoxia pathways in progressing periodontitis.Immunology. 2021 Apr;162(4):405-417. doi: 10.1111/imm.13292. Epub 2021 Jan 4. Immunology. 2021. PMID: 33314069 Free PMC article.
References
-
- Cochran DL. Inflammation and bone loss in periodontal disease. J Periodontol. 2008;79:1569‐1576. - PubMed
-
- Holt SC, Ebersole JL. Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia: the “red complex”, a prototype polybacterial pathogenic consortium in periodontitis. Periodontol 2000. 2005;38:72‐122. - PubMed
-
- Ebersole JL. Humoral immune responses in gingival crevice fluid: local and systemic implications. Periodontol 2000. 2003;31:135‐166. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases