

Aging of spermatogonial stem cells by Jnk-mediated glycolysis activation
- PMID: 31358627
- PMCID: PMC6697785
- DOI: 10.1073/pnas.1904980116
Aging of spermatogonial stem cells by Jnk-mediated glycolysis activation
Abstract
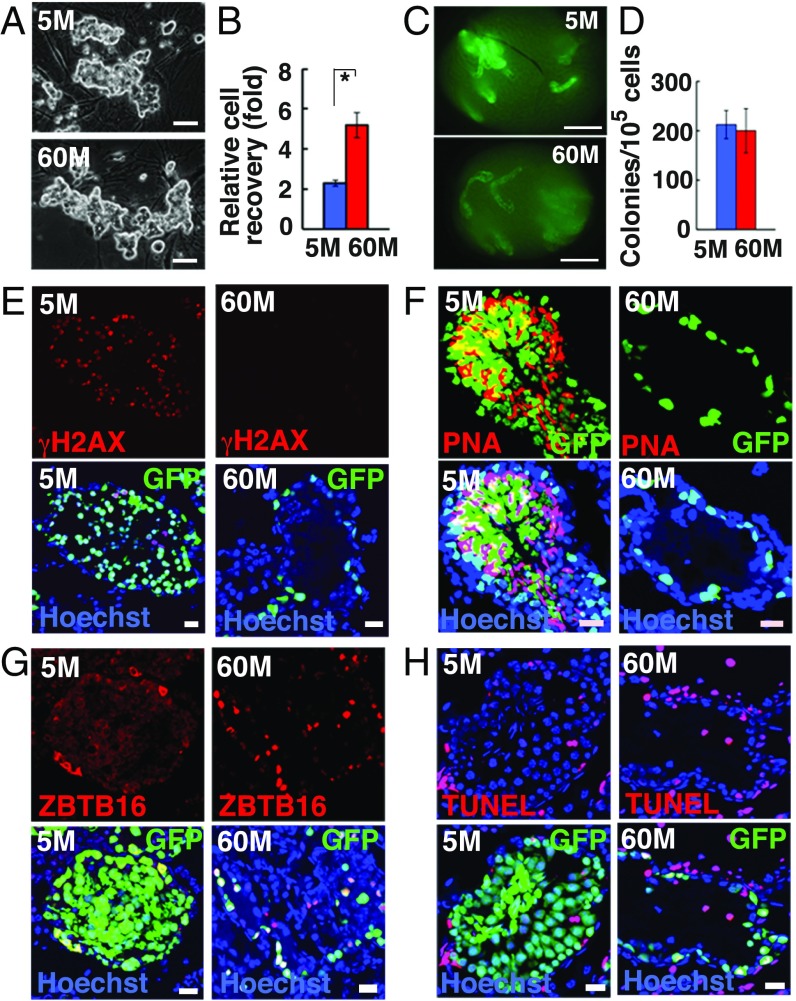
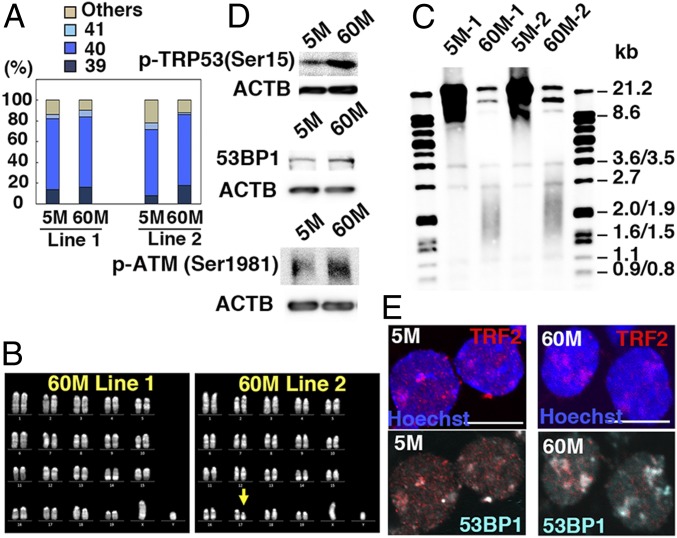
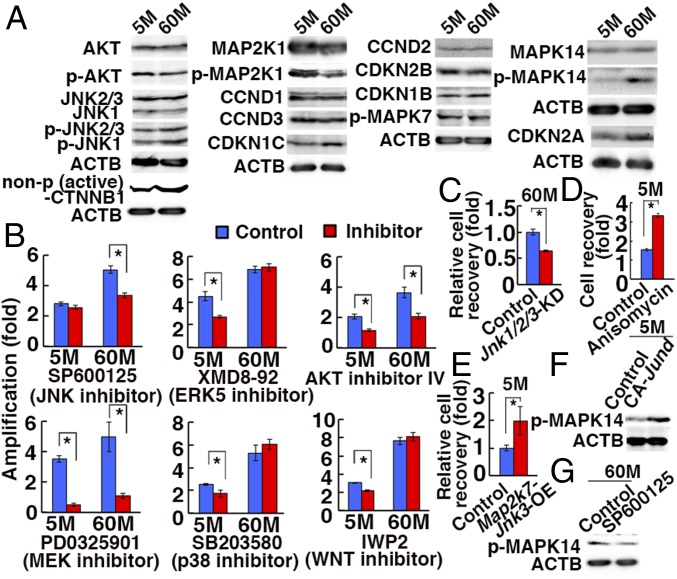
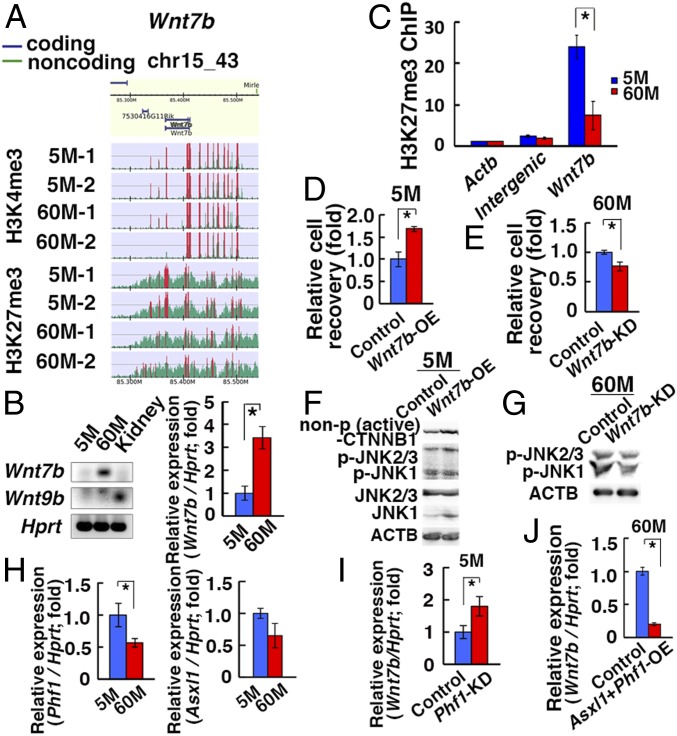
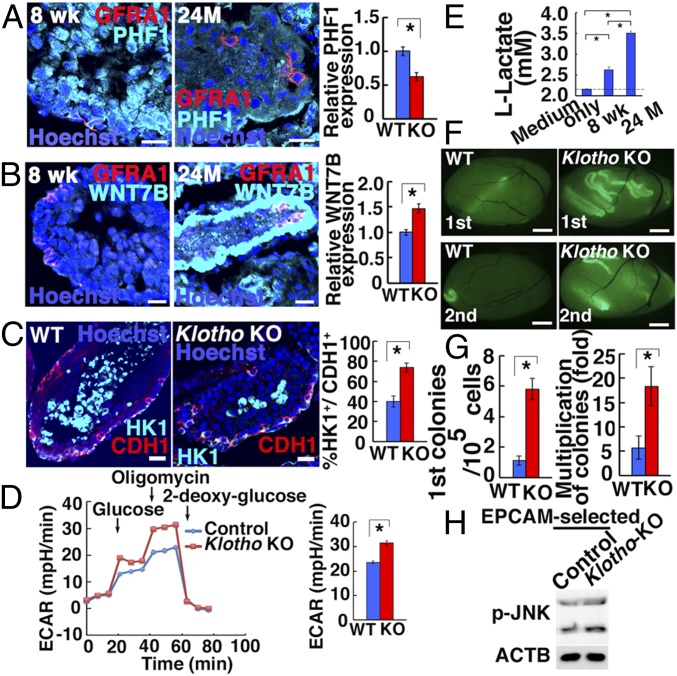
Because spermatogonial stem cells (SSCs) are immortal by serial transplantation, SSC aging in intact testes is considered to be caused by a deteriorated microenvironment. Here, we report a cell-intrinsic mode of SSC aging by glycolysis activation. Using cultured SSCs, we found that aged SSCs proliferated more actively than young SSCs and showed enhanced glycolytic activity. Moreover, they remained euploid and exhibited stable androgenetic imprinting patterns with robust SSC activity despite having shortened telomeres. Aged SSCs showed increased Wnt7b expression, which was associated with decreased Polycomb complex 2 activity. Our results suggest that aberrant Wnt7b expression activated c-jun N-terminal kinase (JNK), which down-regulated mitochondria numbers by suppressing Ppargc1a Down-regulation of Ppargc1a probably decreased reactive oxygen species and enhanced glycolysis. Analyses of the Klotho-deficient aging mouse model and 2-y-old aged rats confirmed JNK hyperactivation and increased glycolysis. Therefore, not only microenvironment but also intrinsic activation of JNK-mediated glycolysis contributes to SSC aging.
Keywords: aging; glycolysis; spermatogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- de Rooij D. G., Russell L. D., All you wanted to know about spermatogonia but were afraid to ask. J. Androl. 21, 776–798 (2000). - PubMed
-
- Meistrich M. L., van Beek M. E. A. B., “Spermatogonial stem cells” in Cell and Molecular Biology of the Testis, Desjardins C. C., Ewing L. L., Eds. (Oxford University Press, 1993), pp. 266–295.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
